

A2 isoform of mammalian translation factor eEF1A displays increased tyrosine phosphorylation and ability to interact with different signalling molecules
- PMID: 17936057
- PMCID: PMC2279807
- DOI: 10.1016/j.biocel.2007.08.014
A2 isoform of mammalian translation factor eEF1A displays increased tyrosine phosphorylation and ability to interact with different signalling molecules
Abstract
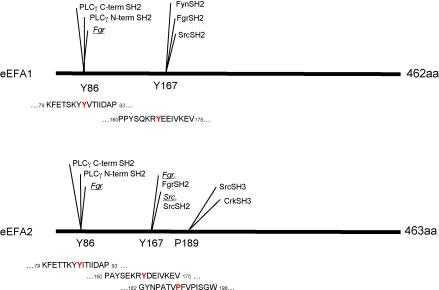
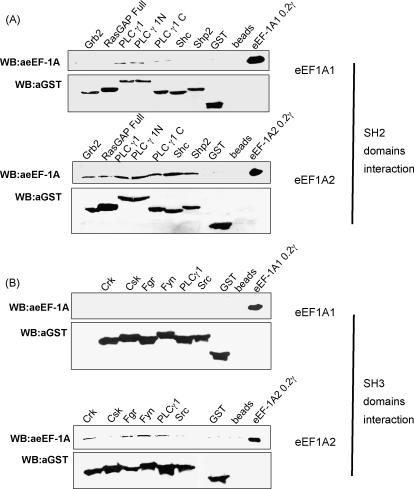
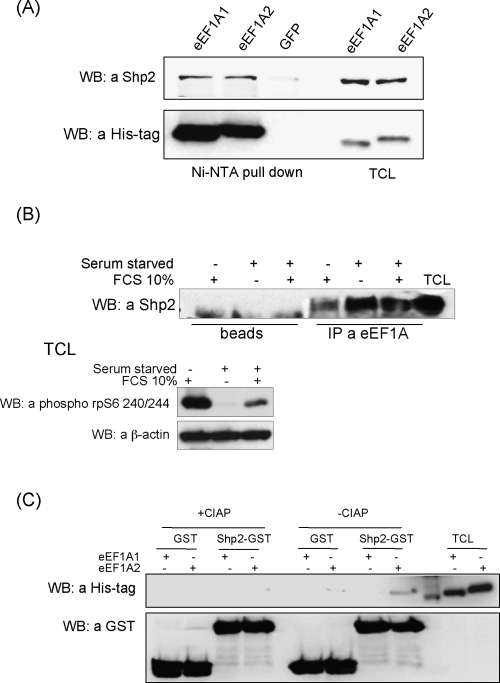
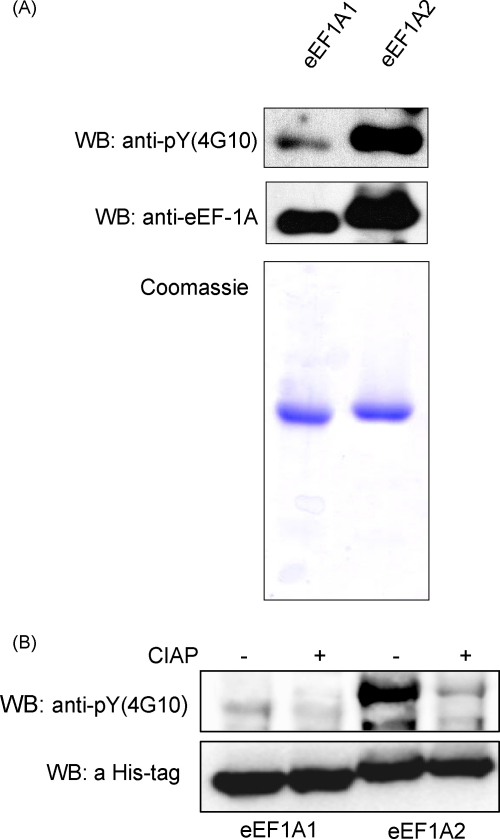
The eEF1A1 and eEF1A2 isoforms of translation elongation factor 1A have 98% similarity and perform the same protein synthesis function catalyzing codon-dependent binding of aminoacyl-tRNA to 80S ribosome. However, the isoforms apparently play different non-canonical roles in apoptosis and cancer development which are awaiting further investigations. We hypothesize that the difference in non-translational functions could be caused, in particular, by differential ability of the isoforms to be involved in phosphotyrosine-mediated signalling. The ability of eEF1A1 and eEF1A2 to interact with SH2 and SH3 domains of different signalling molecules in vitro was compared. Indeed, contrary to eEF1A1, eEF1A2 was able to interact with SH2 domains of Grb2, RasGAP, Shc and C-terminal part of Shp2 as well as with SH3 domains of Crk, Fgr, Fyn and phospholipase C-gamma1. Interestingly, the interaction of both isoforms with Shp2 in vivo was found using stable cell lines expressing eEF1A1-His or eEF1A2-His. The formation of a complex between endogenous eEF1A and Shp2 was also shown. Importantly, a higher level of tyrosine phosphorylation of eEF1A2 as compared to eEF1A1 was demonstrated in several independent experiments and its importance for interaction of eEF1A2 with Shp2 in vitro was revealed. Thus, despite the fact that both isoforms of eEF1A could be involved in the phosphotyrosine-mediated processes, eEF1A2 apparently has greater potential to participate in such signalling pathways. Since tyrosine kinases/phosphatases play a prominent role in human cancerogenesis, our observations may gave a basis for recently found oncogenicity of the eEF1A2 isoform.
Figures
References
-
- Ahmed M., Forsberg J., Bergsten P. Protein profiling of human pancreatic islets by two-dimentional gel electrophoresis and mass-spectrometry. J. Proteome Res. 2005;4(3):931–940. - PubMed
-
- Alper O., Bowden E.T. Novel insights into c-Src. Curr. Pharm. Des. 2005;11(9):1119–1130. - PubMed
-
- Amiri A., Noei F., Jeganathan S., Kulkarni G., Pinke D.E., Lee J.M. eEF1A2 activates Akt and stimulates Akt-dependent actin remodeling, invasion and migration. Oncogene. 2007;26(21):3027–3040. - PubMed
-
- Anand N., Murthy S., Amann G., Wernick M., Porter L.A., Cukier I.H. Protein elongation factor eEF1A2 is a putative oncogene in ovarian cancer. Nat. Genet. 2002;31(3):301–305. - PubMed
-
- Ann D.K., Moutsatsos I.K., Nakamura T., Lin H.H., Mao P.L., Lee M.J. Isolation and characterization of the rat chromosomal gene for a polypeptide (pS1) antigenically related to statin. J. Biol. Chem. 1991;266(16):10429–10437. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
