

Organization of mammalian locomotor rhythm and pattern generation
- PMID: 17936363
- PMCID: PMC2214837
- DOI: 10.1016/j.brainresrev.2007.08.006
Organization of mammalian locomotor rhythm and pattern generation
Abstract
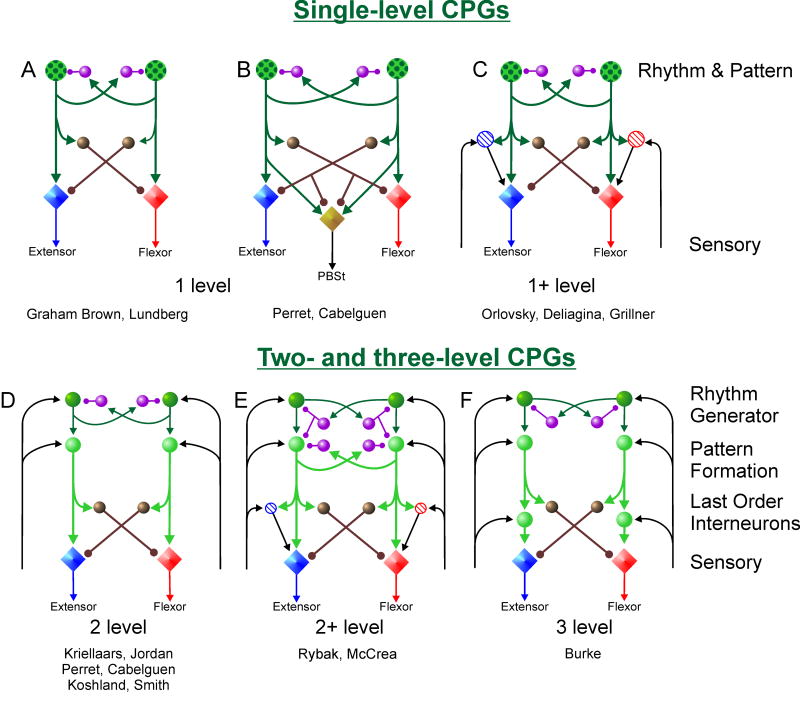
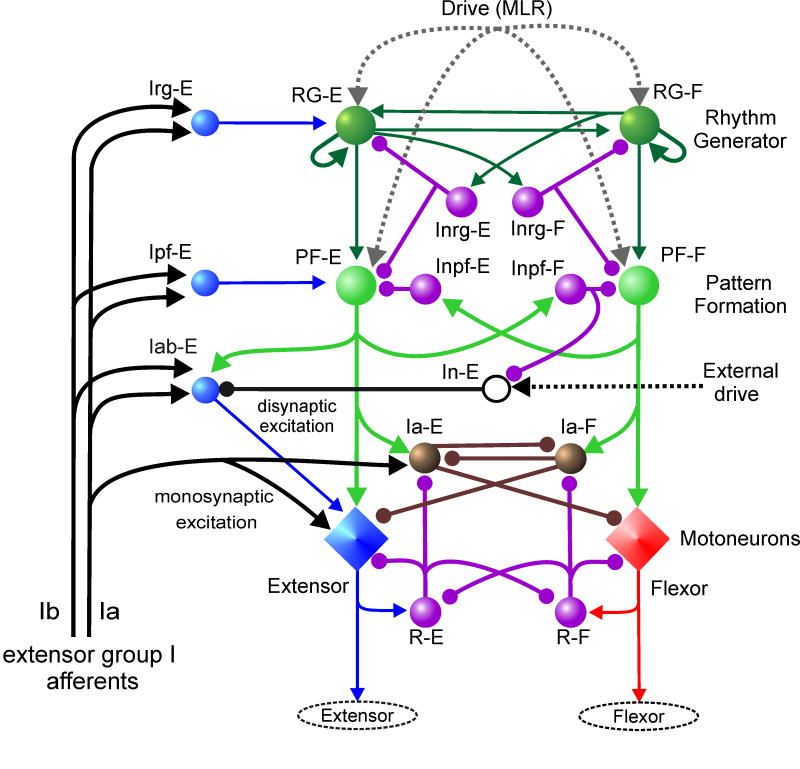
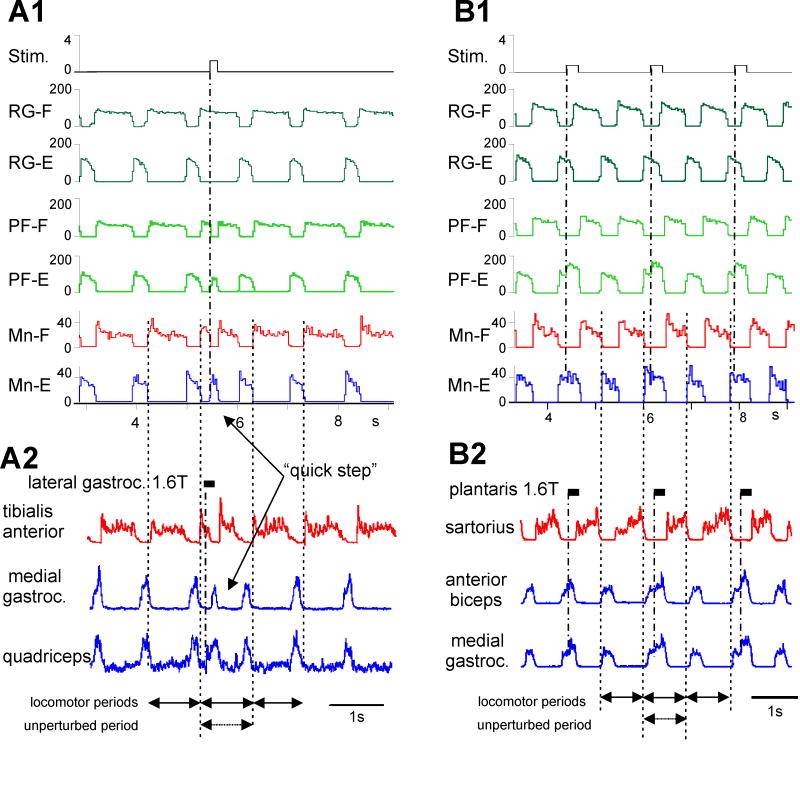
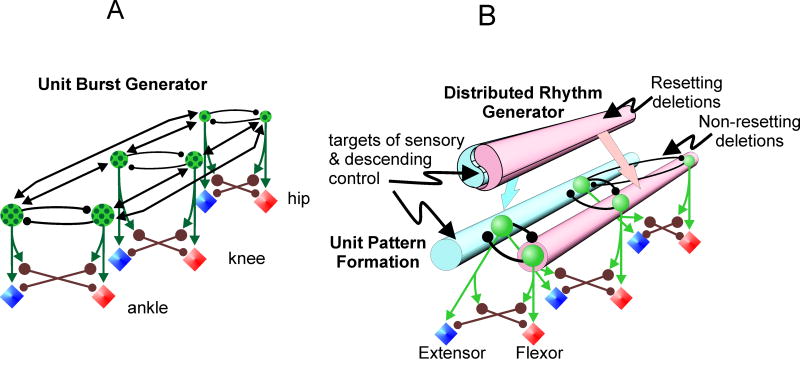
Central pattern generators (CPGs) located in the spinal cord produce the coordinated activation of flexor and extensor motoneurons during locomotion. Previously proposed architectures for the spinal locomotor CPG have included the classical half-center oscillator and the unit burst generator (UBG) comprised of multiple coupled oscillators. We have recently proposed another organization in which a two-level CPG has a common rhythm generator (RG) that controls the operation of the pattern formation (PF) circuitry responsible for motoneuron activation. These architectures are discussed in relation to recent data obtained during fictive locomotion in the decerebrate cat. The data show that the CPG can maintain the period and phase of locomotor oscillations both during spontaneous deletions of motoneuron activity and during sensory stimulation affecting motoneuron activity throughout the limb. The proposed two-level CPG organization has been investigated with a computational model which incorporates interactions between the CPG, spinal circuits and afferent inputs. The model includes interacting populations of spinal interneurons and motoneurons modeled in the Hodgkin-Huxley style. Our simulations demonstrate that a relatively simple CPG with separate RG and PF networks can realistically reproduce many experimental phenomena including spontaneous deletions of motoneuron activity and a variety of effects of afferent stimulation. The model suggests plausible explanations for a number of features of real CPG operation that would be difficult to explain in the framework of the classical single-level CPG organization. Some modeling predictions and directions for further studies of locomotor CPG organization are discussed.
Figures
References
-
- Bernstein N. The coordination and regulation of movements. Pergamon Press; London: 1967.
-
- Bizzi E, Tresch MC, Saltiel P, d’Avella A. New perspectives on spinal motor systems. Nat Rev Neurosci. 2000;1:101–8. - PubMed
-
- Burke RE, Degtyarenko AM, Simon ES. Patterns of locomotor drive to motoneurons and last-order interneurons: clues to the structure of the CPG. J Neurophysiol. 2001;86:447–462. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
