

Wingless activity in the precursor cells specifies neuronal migratory behavior in the Drosophila nerve cord
- PMID: 17936746
- PMCID: PMC2175177
- DOI: 10.1016/j.ydbio.2007.09.004
Wingless activity in the precursor cells specifies neuronal migratory behavior in the Drosophila nerve cord
Abstract
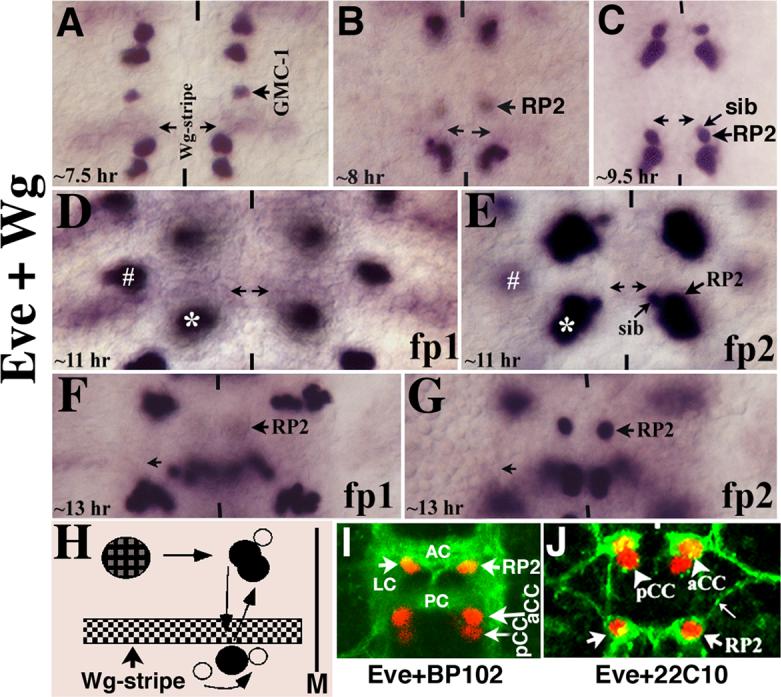
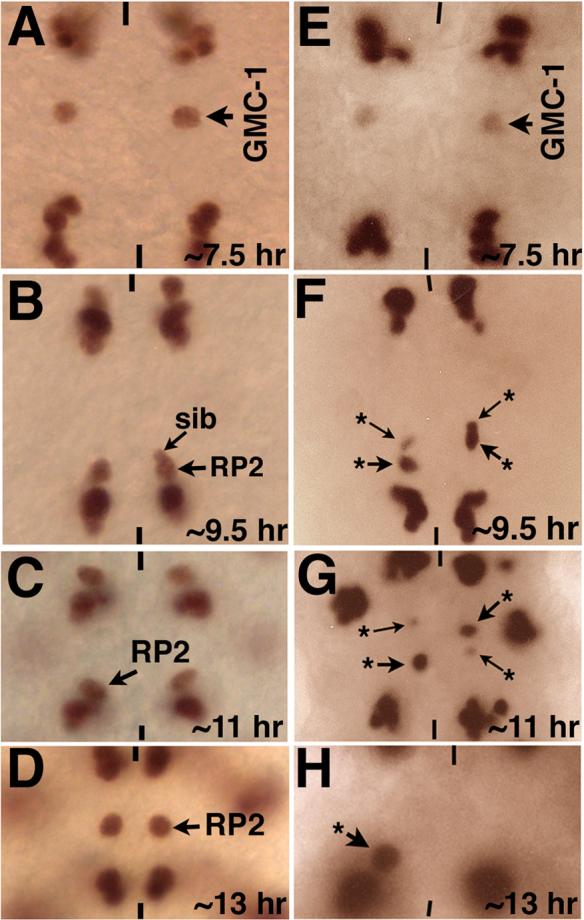
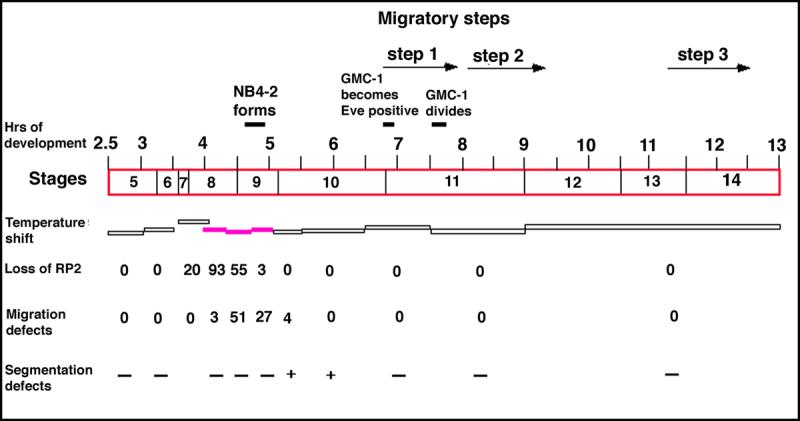
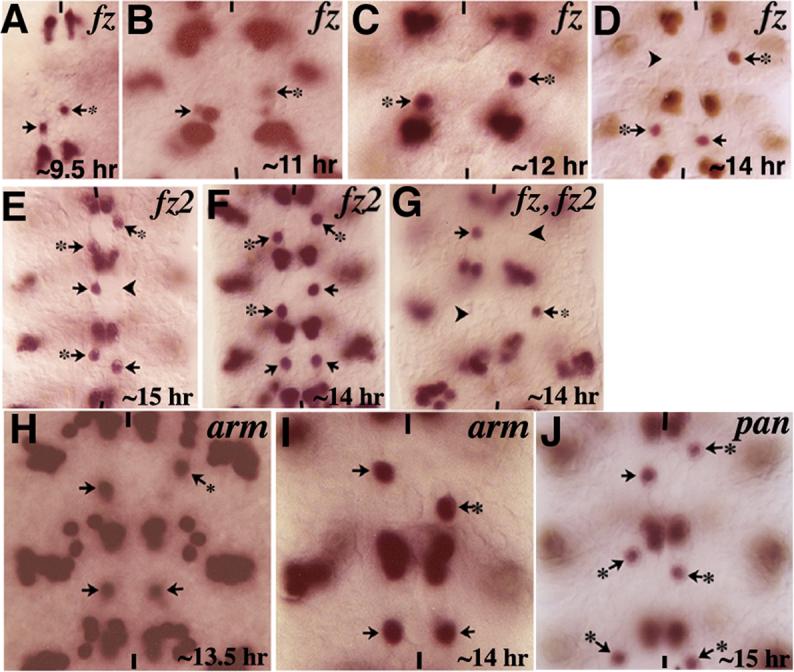
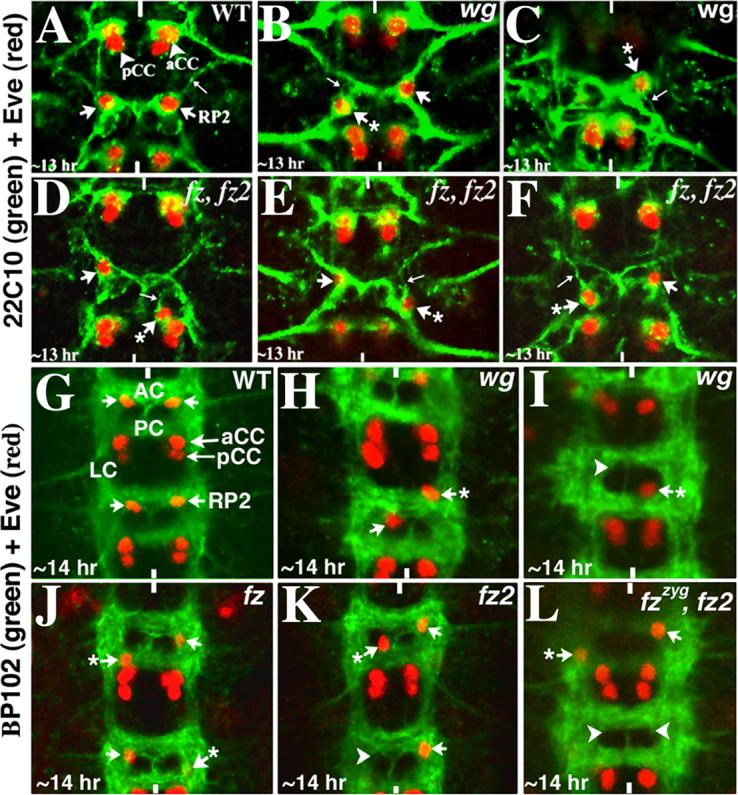
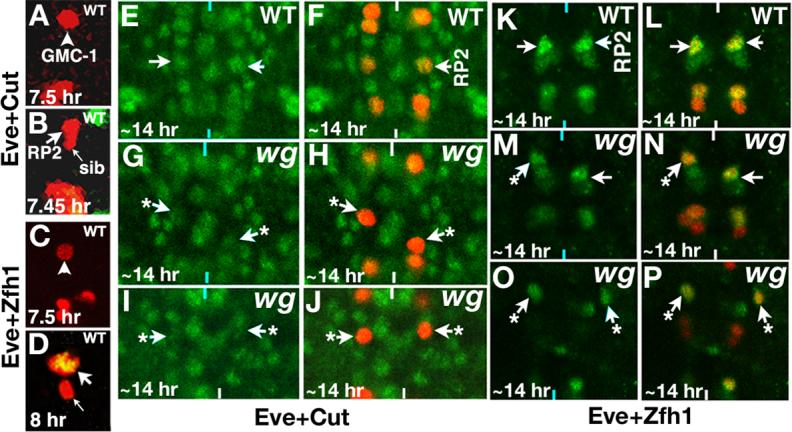
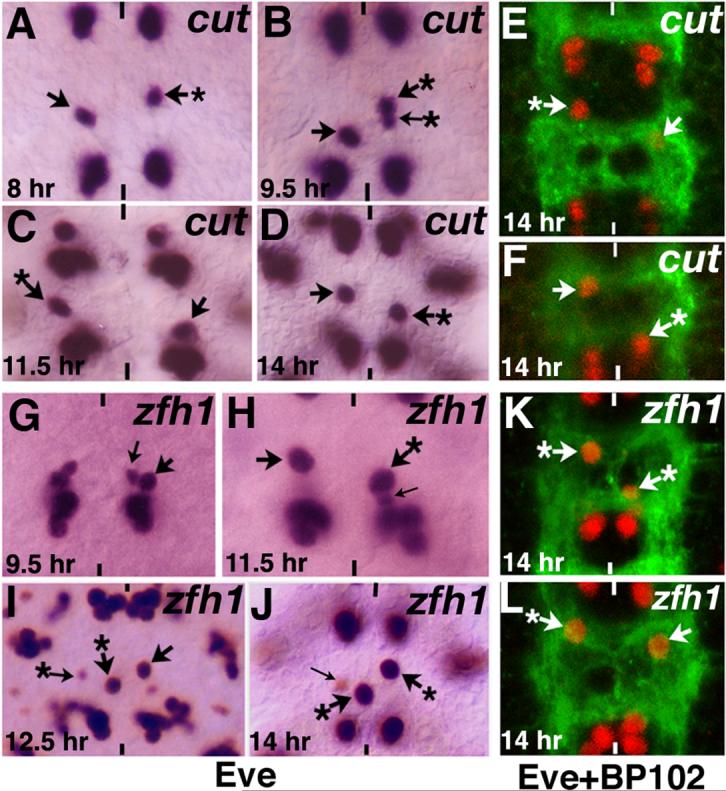
Neurons and their precursor cells are formed in different regions within the developing CNS, but they migrate and occupy very specific sites in the mature CNS. The ultimate position of neurons is crucial for establishing proper synaptic connectivity in the brain. In Drosophila, despite its extensive use as a model system to study neurogenesis, we know almost nothing about neuronal migration or its regulation. In this paper, I show that one of the most studied neuronal pairs in the Drosophila nerve cord, RP2/sib, has a complicated migratory route. Based on my studies on Wingless (Wg) signaling, I report that the neuronal migratory pattern is determined at the precursor cell stage level. The results show that Wg activity in the precursor neuroectodermal and neuroblast levels specify neuronal migratory pattern two divisions later, thus, well ahead of the actual migratory event. Moreover, at least two downstream genes, Cut and Zfh1, are involved in this process but their role is at the downstream neuronal level. The functional importance of normal neuronal migration and the requirement of Wg signaling for the process are indicated by the finding that mislocated RP2 neurons in embryos mutant for Wg-signaling fail to properly send out their axon projection.
Figures
References
-
- Bhanot P, Brink M, Samos CH, Hsieh J-C, Wang Y, Macke JP, Andrew D, Nathans J, Nusse R. A new member of the frizzled family from Drosophila functions as a Wingless receptor. Nature. 1996;382:225–230. - PubMed
-
- Bhat KM. The patched signaling pathway mediates repression of gooseberry allowing neuroblast specification by wingless during Drosophila neurogenesis. Development. 1996;122:2911–2923. - PubMed
-
- Bhat KM. frizzled and frizzled 2 play a partially redundant role in Wingless signaling and have similar requirements to Wingless in neurogenesis. Cell. 1998;95:1027–1036. - PubMed
-
- Bhat KM. Segment polarity genes in neuroblast formation and identity specification during Drosophila neurogenesis. BioEssays. 1999;21:472–485. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
