

Metabolic phenotype of methylmalonic acidemia in mice and humans: the role of skeletal muscle
- PMID: 17937813
- PMCID: PMC2140053
- DOI: 10.1186/1471-2350-8-64
Metabolic phenotype of methylmalonic acidemia in mice and humans: the role of skeletal muscle
Abstract
Background: Mutations in methylmalonyl-CoA mutase cause methylmalonic acidemia, a common organic aciduria. Current treatment regimens rely on dietary management and, in severely affected patients, liver or combined liver-kidney transplantation. For undetermined reasons, transplantation does not correct the biochemical phenotype.
Methods: To study the metabolic disturbances seen in this disorder, we have created a murine model with a null allele at the methylmalonyl-CoA mutase locus and correlated the results observed in the knock-out mice to patient data. To gain insight into the origin and magnitude of methylmalonic acid (MMA) production in humans with methylmalonyl-CoA mutase deficiency, we evaluated two methylmalonic acidemia patients who had received different variants of combined liver-kidney transplants, one with a complete liver replacement-kidney transplant and the other with an auxiliary liver graft-kidney transplant, and compared their metabolite production to four untransplanted patients with intact renal function.
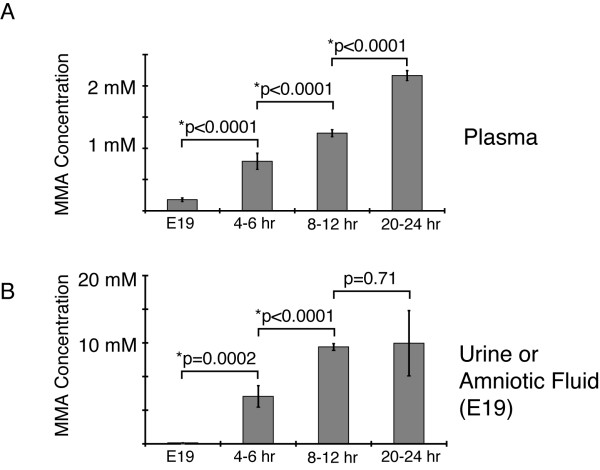
Results: Enzymatic, Western and Northern analyses demonstrated that the targeted allele was null and correctable by lentiviral complementation. Metabolite studies defined the magnitude and tempo of plasma MMA concentrations in the mice. Before a fatal metabolic crisis developed in the first 24-48 hours, the methylmalonic acid content per gram wet-weight was massively elevated in the skeletal muscle as well as the kidneys, liver and brain. Near the end of life, extreme elevations in tissue MMA were present primarily in the liver. The transplant patients studied when well and on dietary therapy, displayed massive elevations of MMA in the plasma and urine, comparable to the levels seen in the untransplanted patients with similar enzymatic phenotypes and dietary regimens.
Conclusion: The combined observations from the murine metabolite studies and patient investigations indicate that during homeostasis, a large portion of circulating MMA has an extra-heptorenal origin and likely derives from the skeletal muscle. Our studies suggest that modulating skeletal muscle metabolism may represent a strategy to increase metabolic capacity in methylmalonic acidemia as well as other organic acidurias. This mouse model will be useful for further investigations exploring disease mechanisms and therapeutic interventions in methylmalonic acidemia, a devastating disorder of intermediary metabolism.
Figures




Similar articles
-
Adenoviral-mediated correction of methylmalonyl-CoA mutase deficiency in murine fibroblasts and human hepatocytes.BMC Med Genet. 2007 Apr 30;8:24. doi: 10.1186/1471-2350-8-24. BMC Med Genet. 2007. PMID: 17470278 Free PMC article.
-
Adenovirus-mediated gene delivery rescues a neonatal lethal murine model of mut(0) methylmalonic acidemia.Hum Gene Ther. 2008 Jan;19(1):53-60. doi: 10.1089/hum.2007.0118. Hum Gene Ther. 2008. PMID: 18052792 Free PMC article.
-
Renal transplant in methylmalonic acidemia: could it be the best option? Report on a case at 10 years and review of the literature.Pediatr Nephrol. 2007 Aug;22(8):1209-14. doi: 10.1007/s00467-007-0460-z. Epub 2007 Mar 31. Pediatr Nephrol. 2007. PMID: 17401587 Review.
-
A detailed analysis of methylmalonic acid kinetics during hemodialysis and after combined liver/kidney transplantation in a patient with mut (0) methylmalonic acidemia.J Inherit Metab Dis. 2014 Nov;37(6):899-907. doi: 10.1007/s10545-014-9730-7. Epub 2014 Jun 25. J Inherit Metab Dis. 2014. PMID: 24961826 Free PMC article.
-
Systematic literature review and meta-analysis on the epidemiology of methylmalonic acidemia (MMA) with a focus on MMA caused by methylmalonyl-CoA mutase (mut) deficiency.Orphanet J Rare Dis. 2019 Apr 25;14(1):84. doi: 10.1186/s13023-019-1063-z. Orphanet J Rare Dis. 2019. PMID: 31023387 Free PMC article.
Cited by
-
Liver-directed recombinant adeno-associated viral gene delivery rescues a lethal mouse model of methylmalonic acidemia and provides long-term phenotypic correction.Hum Gene Ther. 2010 Sep;21(9):1147-54. doi: 10.1089/hum.2010.008. Hum Gene Ther. 2010. PMID: 20486773 Free PMC article.
-
Gene therapy in a murine model of methylmalonic acidemia using rAAV9-mediated gene delivery.Gene Ther. 2012 Apr;19(4):385-91. doi: 10.1038/gt.2011.108. Epub 2011 Jul 21. Gene Ther. 2012. PMID: 21776024 Free PMC article.
-
Systemic gene therapy for methylmalonic acidemia using the novel adeno-associated viral vector 44.9.Mol Ther Methods Clin Dev. 2022 Sep 6;27:61-72. doi: 10.1016/j.omtm.2022.09.001. eCollection 2022 Dec 8. Mol Ther Methods Clin Dev. 2022. PMID: 36186952 Free PMC article.
-
Defects in muscle branched-chain amino acid oxidation contribute to impaired lipid metabolism.Mol Metab. 2016 Aug 6;5(10):926-936. doi: 10.1016/j.molmet.2016.08.001. eCollection 2016 Oct. Mol Metab. 2016. PMID: 27689005 Free PMC article.
-
Biomarkers to predict disease progression and therapeutic response in isolated methylmalonic acidemia.J Inherit Metab Dis. 2023 Jul;46(4):554-572. doi: 10.1002/jimd.12636. Epub 2023 Jun 6. J Inherit Metab Dis. 2023. PMID: 37243446 Free PMC article. Review.
References
-
- Fenton WA, Gravel RA, Rosenblatt DS. In: Disorders of Propionate and Methylmalonate Metabolism in The Metabolic and Molecular Bases for Inhertited Disease. 8. Scriver CR, Beaudet AL, Sly WS, Valle D, editor. McGraw-Hill, Inc., New York; 2001. pp. 2165–2192.
-
- Fenton WA, Rosenblatt DS. In: Inherited Disorders of Folate and Cobalamin Transport and Metabolism in The Metabolic and Molecular Bases of Inherited Disease. 8. Scriver CR BA, Sly WS, Valle D, Childs B, Kinzler KW, Vogelstein B, editor. McGraw-Hill; 2001. pp. 3897–3933.
-
- Matsui SM, Mahoney MJ, Rosenberg LE. The natural history of the inherited methylmalonic acidemias. N Engl J Med. 1983;308:857–861. - PubMed
-
- van der Meer SB, Poggi F, Spada M, Bonnefont JP, Ogier H, Hubert P, Depondt E, Rapoport D, Rabier D, Charpentier C, et al. Clinical outcome of long-term management of patients with vitamin B12-unresponsive methylmalonic acidemia. J Pediatr. 1994;125:903–908. doi: 10.1016/S0022-3476(05)82005-0. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
