

Impairment of infectivity and immunoprotective effect of a LYT1 null mutant of Trypanosoma cruzi
- PMID: 17938222
- PMCID: PMC2223657
- DOI: 10.1128/IAI.00400-07
Impairment of infectivity and immunoprotective effect of a LYT1 null mutant of Trypanosoma cruzi
Abstract
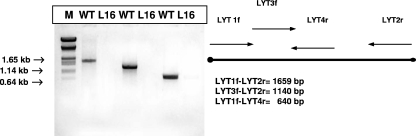
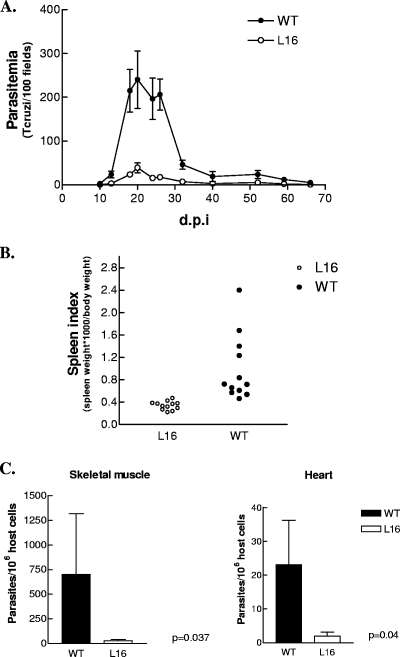
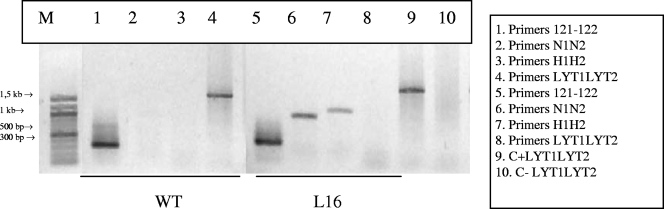
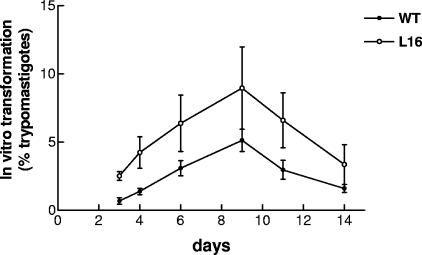
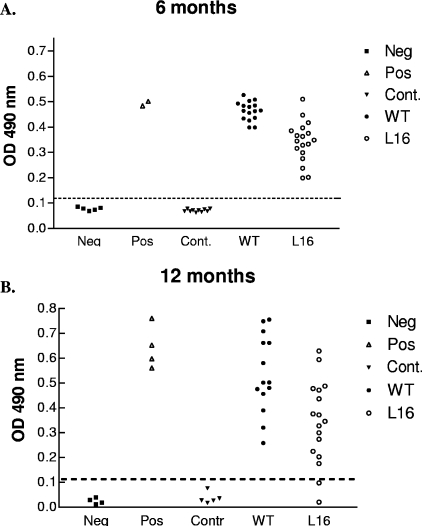
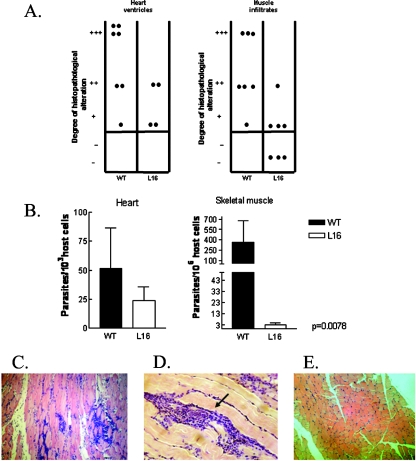
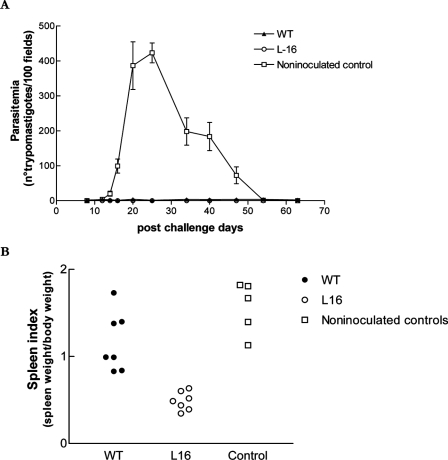
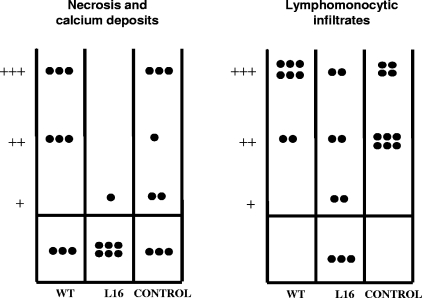
Trypanosoma cruzi infection of host cells is a complex process in which many proteins participate but only a few of these proteins have been identified experimentally. One parasite factor likely to be involved is the protein product of LYT1, a single-copy gene cloned, sequenced, and characterized by Manning-Cela et al. (Infect. Immun. 69:3916-3923, 2001). This gene was potentially associated with infectivity, since the deletion of both LYT1 alleles in the CL Brenner strain (the wild type [WT]) resulted in a null mutant T. cruzi clone (L16) that shows an attenuated phenotype in cell culture models. The aim of this work was to characterize the infective behavior of L16 in the insect vector and murine models. The infection of adult Swiss mice with 10(3) trypomastigotes of both clones revealed a significant reduction in infective behavior of L16, as shown by direct parasitemia, spleen index, and quantitation of tissue parasite burden, suggesting the loss of virulence in the null mutant clone. Although L16 blood counts were almost undetectable, blood-based PCRs indicated the presence of latent and persistent infection during all of the study period and epimastigotes were reisolated from hemocultures until 12 months postinfection. Nevertheless, virulence was not restored in L16 by serial passages in mice, and reisolated parasites lacking the LYT1 gene and bearing the antibiotic resistance genes revealed the stability of the genetic manipulation. Histopathological studies showed a strong diminution in the muscle inflammatory response triggered by L16 compared to that triggered by the WT group, consistent with a lower tissue parasite load. A strong protection against a virulent challenge in both L16- and WT-infected mice was observed; however, the immunizing infection by the genetically modified parasite was highly attenuated.
Figures
Similar articles
-
Knockout of the dhfr-ts gene in Trypanosoma cruzi generates attenuated parasites able to confer protection against a virulent challenge.PLoS Negl Trop Dis. 2011 Dec;5(12):e1418. doi: 10.1371/journal.pntd.0001418. Epub 2011 Dec 13. PLoS Negl Trop Dis. 2011. PMID: 22180798 Free PMC article.
-
Targeted deletion of the gp72 gene decreases the infectivity of Trypanosoma cruzi for mice and insect vectors.J Parasitol. 2002 Jun;88(3):489-93. doi: 10.1645/0022-3395(2002)088[0489:TDOTGG]2.0.CO;2. J Parasitol. 2002. PMID: 12099416
-
Trypanosoma cruzi carrying a targeted deletion of a Tc52 protein-encoding allele elicits attenuated Chagas' disease in mice.Immunol Lett. 2003 Oct 9;89(1):67-80. doi: 10.1016/s0165-2478(03)00112-3. Immunol Lett. 2003. PMID: 12946866
-
Biological characterization of Trypanosoma cruzi stocks from Chilean insect vectors.Exp Parasitol. 1998 Jul;89(3):312-22. doi: 10.1006/expr.1998.4289. Exp Parasitol. 1998. PMID: 9676709
-
Role of Virulence Factors of Trypanosomatids in the Insect Vector and Putative Genetic Events Involved in Surface Protein Diversity.Front Cell Infect Microbiol. 2022 Apr 28;12:807172. doi: 10.3389/fcimb.2022.807172. eCollection 2022. Front Cell Infect Microbiol. 2022. PMID: 35573777 Free PMC article. Review.
Cited by
-
A monoallelic deletion of the TcCRT gene increases the attenuation of a cultured Trypanosoma cruzi strain, protecting against an in vivo virulent challenge.PLoS Negl Trop Dis. 2014 Feb 13;8(2):e2696. doi: 10.1371/journal.pntd.0002696. eCollection 2014 Feb. PLoS Negl Trop Dis. 2014. PMID: 24551259 Free PMC article.
-
Knockout of the dhfr-ts gene in Trypanosoma cruzi generates attenuated parasites able to confer protection against a virulent challenge.PLoS Negl Trop Dis. 2011 Dec;5(12):e1418. doi: 10.1371/journal.pntd.0001418. Epub 2011 Dec 13. PLoS Negl Trop Dis. 2011. PMID: 22180798 Free PMC article.
-
Immunization with a Trypanosoma cruzi cyclophilin-19 deletion mutant protects against acute Chagas disease in mice.NPJ Vaccines. 2023 Apr 25;8(1):63. doi: 10.1038/s41541-023-00647-5. NPJ Vaccines. 2023. PMID: 37185599 Free PMC article.
-
Genetically attenuated Trypanosoma cruzi parasites as a potential vaccination tool.Bioengineered. 2012 Jul-Aug;3(4):242-6. doi: 10.4161/bioe.20680. Epub 2012 Jun 18. Bioengineered. 2012. PMID: 22705838 Free PMC article.
-
Molecular mechanisms of host cell invasion by Trypanosoma cruzi.Exp Parasitol. 2010 Nov;126(3):283-91. doi: 10.1016/j.exppara.2010.06.023. Epub 2010 Jun 18. Exp Parasitol. 2010. PMID: 20599990 Free PMC article. Review.
References
-
- Allaoui, A., C. Francois, K. Zemzoumi, E. Guilvard, and A. Ouaissi. 1999. Intracellular growth and metacyclogenesis defects in Trypanosoma cruzi carrying a targeted deletion of a Tc52 protein-encoding allele. Mol. Microbiol. 321273-1286. - PubMed
-
- Andrade, L. O., and N. W. Andrews. 2005. The Trypanosoma cruzi-host-cell interplay: location, invasion, retention. Nat. Rev. Microbiol. 3819-823. - PubMed
-
- Andrews, N. W. 1993. Living dangerously: how Trypanosoma cruzi uses lysosomes to get inside host cells and then escapes into the cytoplasm. Biol. Res. 2665-67. - PubMed
-
- Andrews, N. W., C. K. Abrams, S. L. Slatin, and G. Griffiths. 1990. A T. cruzi-secreted protein immunologically related to the complement component C9: evidence for membrane pore-forming activity at low pH. Cell 611277-1287. - PubMed
-
- Basombrio, M. A. 1981. Resistance to re-infection by Trypanosoma cruzi in Chagasic mice. Medicina (B Aires) 41(Suppl.)230-232. (In Spanish.) - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
