

Polyphosphate kinase 1, a conserved bacterial enzyme, in a eukaryote, Dictyostelium discoideum, with a role in cytokinesis
- PMID: 17940044
- PMCID: PMC2034253
- DOI: 10.1073/pnas.0706847104
Polyphosphate kinase 1, a conserved bacterial enzyme, in a eukaryote, Dictyostelium discoideum, with a role in cytokinesis
Abstract
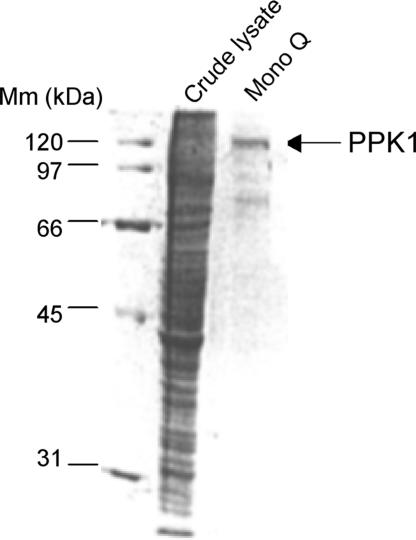
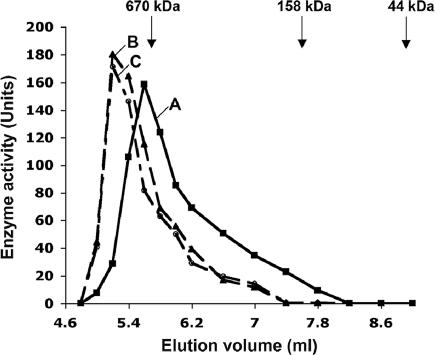
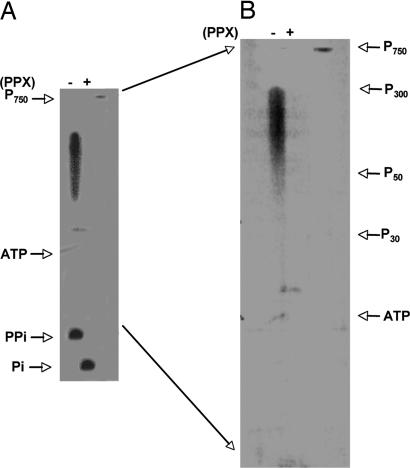
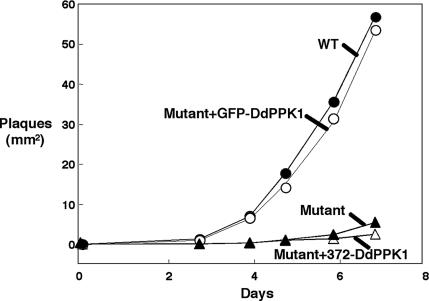
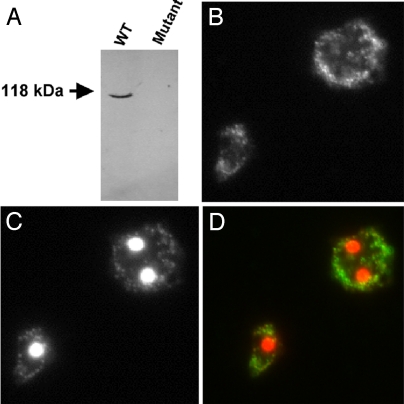
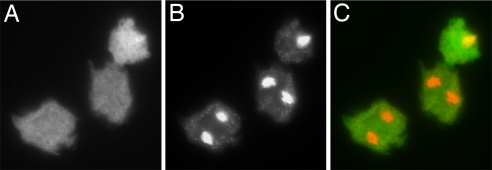
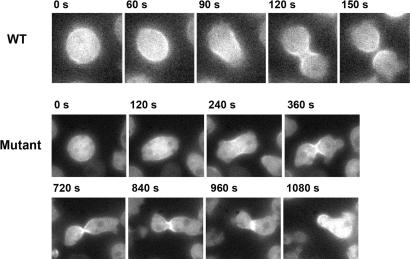
Polyphosphate kinase 1 (PPK1), the principal enzyme responsible for reversible synthesis of polyphosphate (poly P) from the terminal phosphate of ATP, is highly conserved in bacteria and archaea. Dictyostelium discoideum, a social slime mold, is one of a few eukaryotes known to possess a PPK1 homolog (DdPPK1). Compared with PPK1 of Escherichia coli, DdPPK1 contains the conserved residues for ATP binding and autophosphorylation, but has an N-terminal extension of 370 aa, lacking homology with any known protein. Polyphosphate or ATP promote oligomerization of the enzyme in vitro. The DdPPK1 products are heterogeneous in chain length and shorter than those of E. coli. The unique DdPPK1 N-terminal domain was shown to be necessary for its enzymatic activity, cellular localization, and physiological functions. Mutants of DdPPK1, as previously reported, are defective in development, sporulation, and predation, and as shown here, in late stages of cytokinesis and cell division.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
