

Control of corolla monosymmetry in the Brassicaceae Iberis amara
- PMID: 17940055
- PMCID: PMC2034219
- DOI: 10.1073/pnas.0705338104
Control of corolla monosymmetry in the Brassicaceae Iberis amara
Abstract
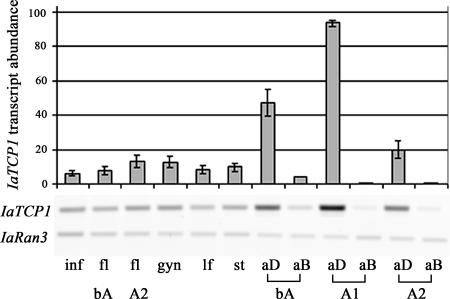
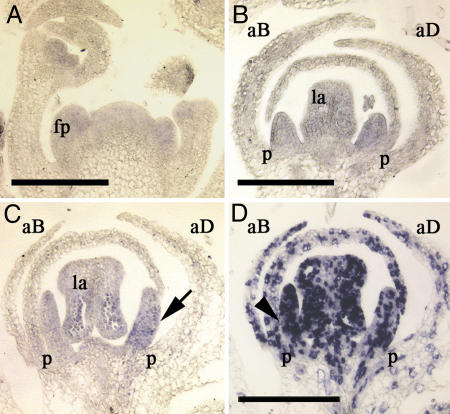
Establishment of morphological novelties has contributed to the enormous diversification of floral architecture. One such novelty, flower monosymmetry, is assumed to have evolved several times independently during angiosperm evolution. To date, analysis of monosymmetry regulation has focused on species from taxa where monosymmetry prevails, such as the Lamiales and Fabaceae. In Antirrhinum majus, formation of a monosymmetric corolla is specified by the activity of the TCP transcription factors CYCLOIDEA (CYC) and DICHOTOMA (DICH). It was shown that establishment of monosymmetry likely requires an early asymmetric floral expression of CYC homologs that needs to be maintained until late floral stages. To understand how CYC homologs might have been recruited during evolution to establish monosymmetry, we characterized the likely CYC ortholog IaTCP1 from Iberis amara (Brassicaceae). Species of the genus Iberis form a monosymmetric corolla, whereas the Brassicaceae are otherwise dominated by genera developing a polysymmetric corolla. Instead of four equally sized petals, I. amara produces two small adaxial and two large abaxial petals. The timing of IaTCP1 expression differs from that of its Arabidopsis homolog TCP1 and other CYC homologs. IaTCP1 lacks an asymmetric early expression but displays a very strong differential expression in the corolla at later floral stages, when the strongest unequal petal growth occurs. Analysis of occasionally occurring peloric Iberis flower variants and comparative functional studies of TCP homologs in Arabidopsis demonstrate the importance of an altered temporal IaTCP1 expression within the Brassicaceae to govern the formation of a monosymmetric corolla.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
