

Optimally wired subnetwork determines neuroanatomy of Caenorhabditis elegans
- PMID: 17942697
- PMCID: PMC2040432
- DOI: 10.1073/pnas.0703183104
Optimally wired subnetwork determines neuroanatomy of Caenorhabditis elegans
Abstract
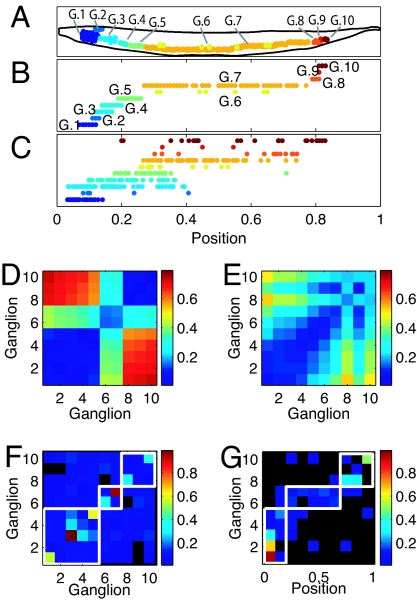
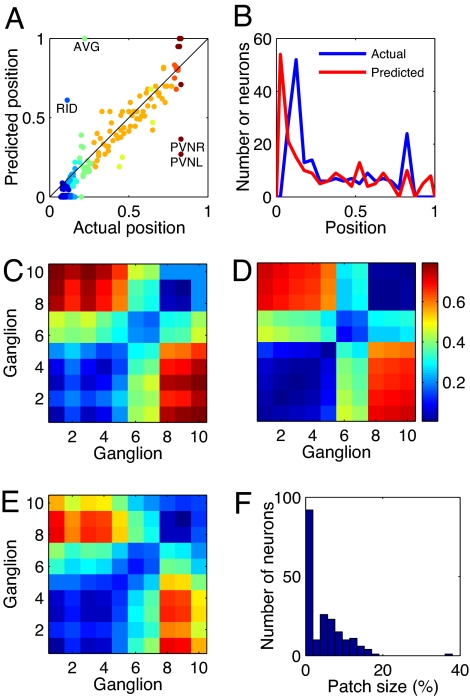
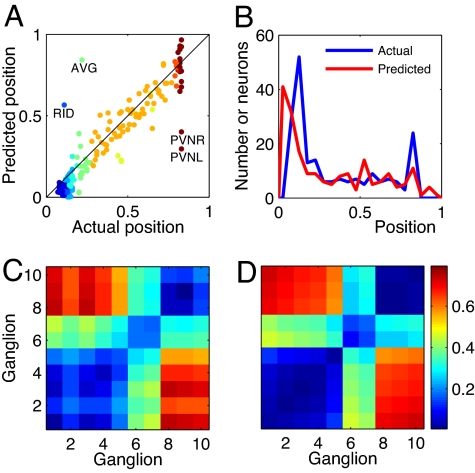
Wiring cost minimization has successfully explained many structures of nervous systems. However, in the nematode Caenorhabditis elegans, for which anatomical data are most detailed, wiring economy is thought to play only a partial role and alone has failed to account for the grouping of neurons into ganglia [Chen BL, Hall DH, Chklovskii DB (2006) Proc Natl Acad Sci USA 103:4723-4728; Kaiser M, Hilgetag CC (2006) PLoS Comput Biol 2:e95; Ahn Y-Y, Jeong H, Kim BJ (2006) Physica A 367:531-537]. Here, we test the hypothesis that optimally wired subnetworks can exist within nonoptimal networks, thus allowing wiring economy to give an improved prediction of spatial structure. We show in C. elegans that the small subnetwork of wires connecting sensory and motor neurons with sensors and muscles, comprising only 15% of connections, is close to optimal and alone predicts the main features of the spatial segregation of neurons into ganglia and encephalization. Moreover, a method to dissect networks into optimal and nonoptimal components is shown to find a large near-optimal subnetwork of 84% of neurons with a very low position error of 5.4%, and that explains clustering of neurons into ganglia and encephalization to fine detail. In general, we expect realistic networks not to be globally optimal in wire cost. We thus propose the strategy of using near-optimal subnetworks to understand neuroanatomical structure.
Conflict of interest statement
The authors declare no conflict of interest.
Figures


References
-
- Striedter GF. Principles of Brain Evolution. Sunderland, MA: Sinauer; 2004.
-
- Nusbaum MP, Blitz DM, Swensen AM, Wood D, Marder E. Trends Neurosci. 2001;24:146–154. - PubMed
-
- Konishi M. In: Elsevier's Encyclopedia of Neuroscience. Adelman G, Smith BH, editors. Amsterdam: Elsevier; 1999. pp. 1906–1908.
-
- Ramón y, Cajal S. Textura del Sistema Nervioso del Hombre y de los Vertebrados. Madrid: Nicolás Moya; 1899. Spanish. trans Pasik P, Pasik T (1999) Texture of the Nervous System of Man and the Vertebrates (Springer, New York)
-
- Mitchison G. Proc R Soc London B Biol Sci. 1991;245:151–158. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Miscellaneous
