

MBNL binds similar RNA structures in the CUG repeats of myotonic dystrophy and its pre-mRNA substrate cardiac troponin T
- PMID: 17942744
- PMCID: PMC2080590
- DOI: 10.1261/rna.610607
MBNL binds similar RNA structures in the CUG repeats of myotonic dystrophy and its pre-mRNA substrate cardiac troponin T
Abstract
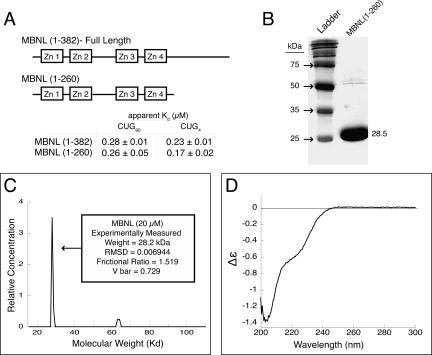
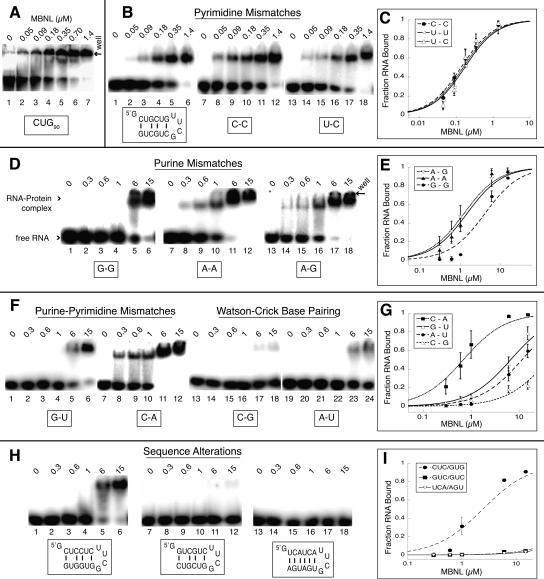
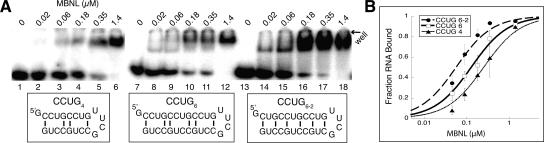
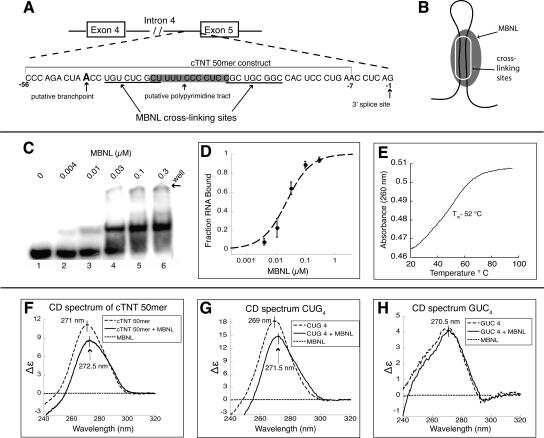
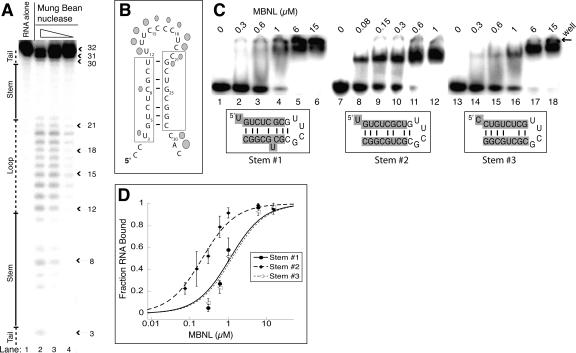
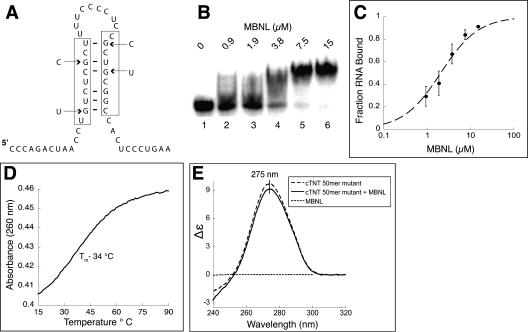
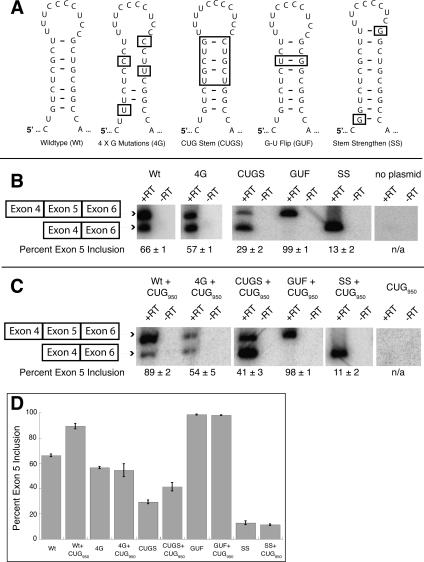
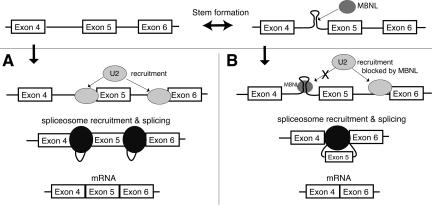
Myotonic dystrophy (DM) is a genetic disorder with multisystemic symptoms that is caused by expression (as RNA) of expanded repeats of CTG or CCTG in the genome. It is hypothesized that the RNA splicing factor muscleblind-like (MBNL) is sequestered to the expanded CUG or CCUG RNAs. Mislocalization of MBNL results in missplicing of a subset of pre-mRNAs that are linked to the symptoms found in DM patients. We demonstrate that MBNL can bind short structured CUG and CCUG repeats with high affinity and specificity. Only 6 base pairs are necessary for MBNL binding: two pyrimidine mismatches and four guanosine-cytosine base pairs in a stem. MBNL also has a preference for pyrimidine mismatches, but many other mismatches are tolerated with decreased affinity. We also demonstrate that MBNL binds the helical region of a stem-loop in the endogenous pre-mRNA target, the cardiac troponin T (cTNT) pre-mRNA. The stem-loop contains two mismatches and resembles both CUG and CCUG repeats. In vivo splicing results indicate that MBNL-regulated splicing is dependent upon the formation of stem-loops recognized by MBNL. These results suggest that MBNL may bind all of its RNA substrates, both normal and pathogenic, as structured stem-loops containing pyrimidine mismatches.
Figures
References
-
- Begemann, G., Paricio, N., Artero, R., Kiss, I., Perez-Alonso, M., Mlodzik, M. Muscleblind, a gene required for photoreceptor differentiation in Drosophila, encodes novel nuclear Cys3His-type zinc-finger-containing proteins. Development. 1997;124:4321–4331. - PubMed
-
- Blencowe, B.J., Bowman, J.A., McCracken, S., Rosonina, E. SR-related proteins and the processing of messenger RNA precursors. Biochem. Cell Biol. 1999;77:277–291. - PubMed
-
- Brown, R.S. Zinc finger proteins: Getting a grip on RNA. Curr. Opin. Struct. Biol. 2005;15:94–98. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous