

Local and global structure of the monomeric subunit of the potassium channel KcsA probed by NMR
- PMID: 17945182
- PMCID: PMC2174260
- DOI: 10.1016/j.bbamem.2007.08.006
Local and global structure of the monomeric subunit of the potassium channel KcsA probed by NMR
Abstract
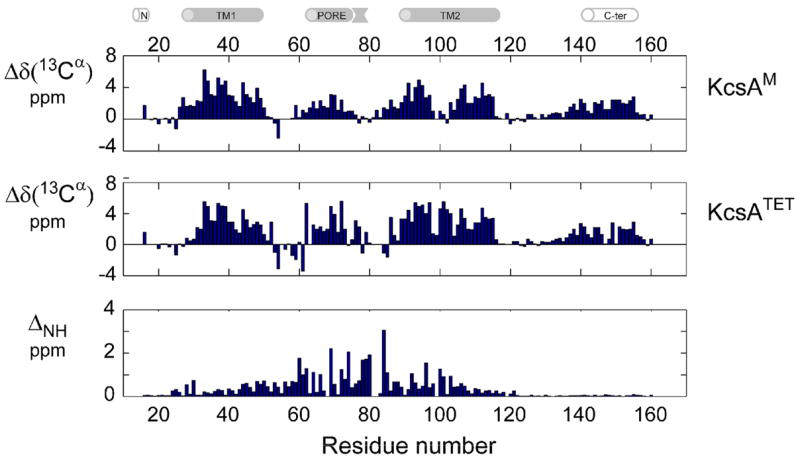
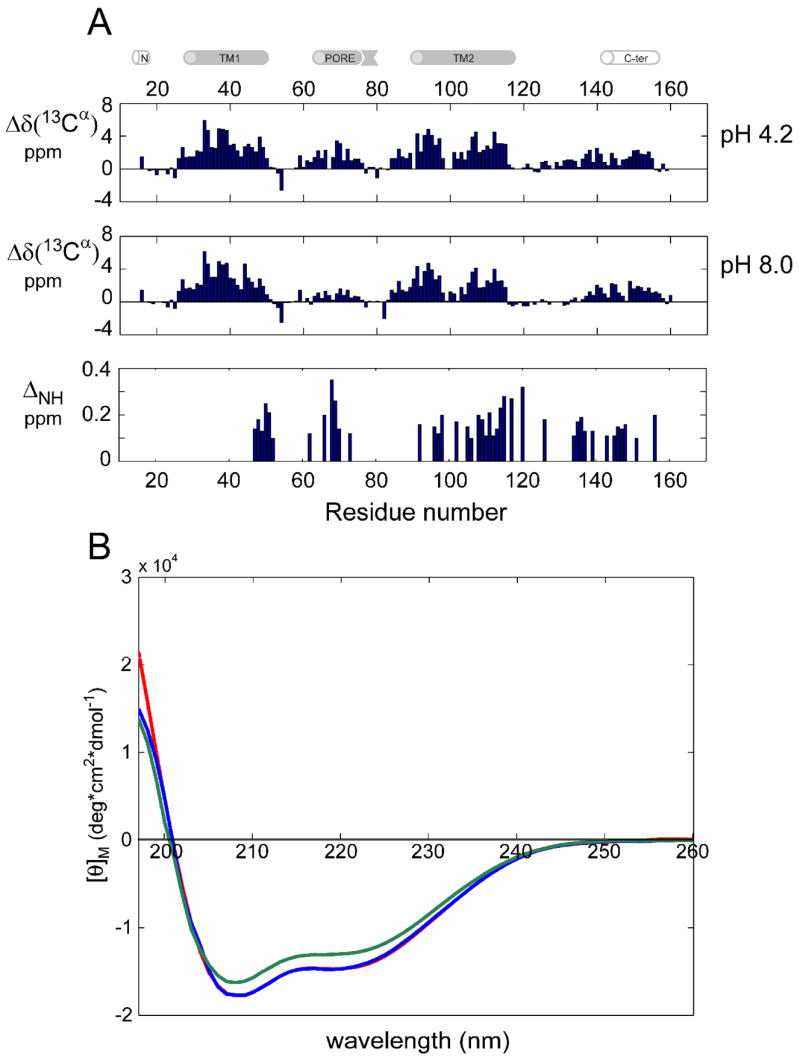
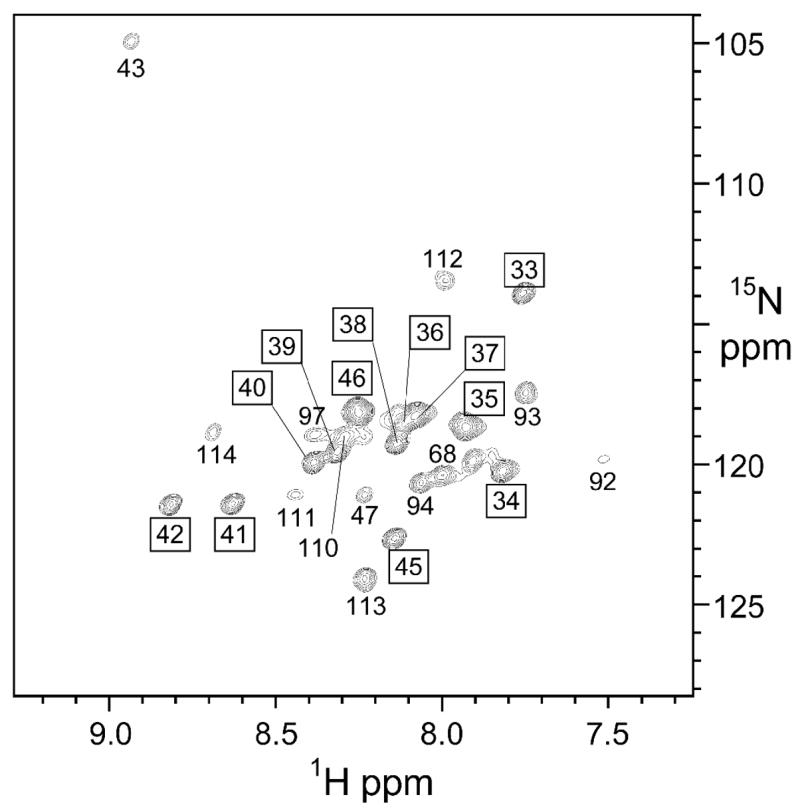
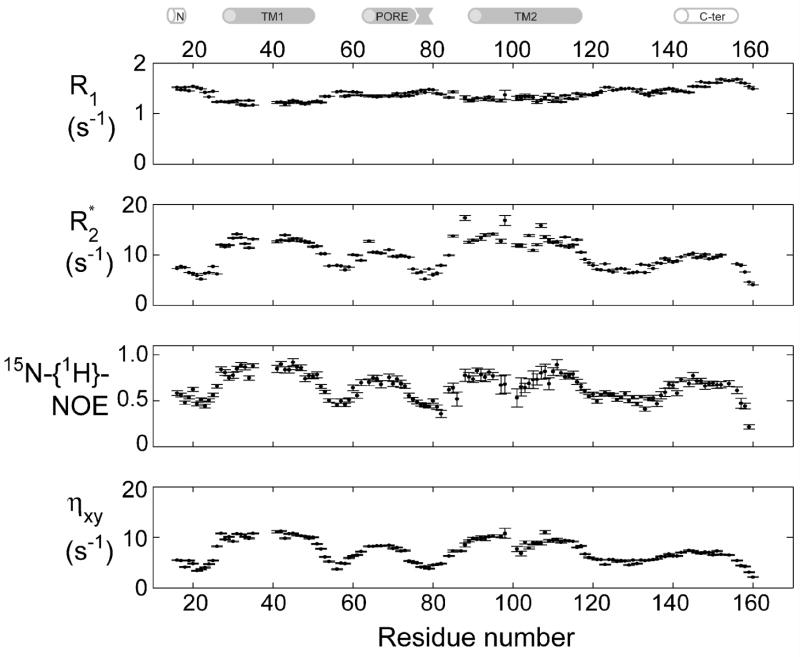
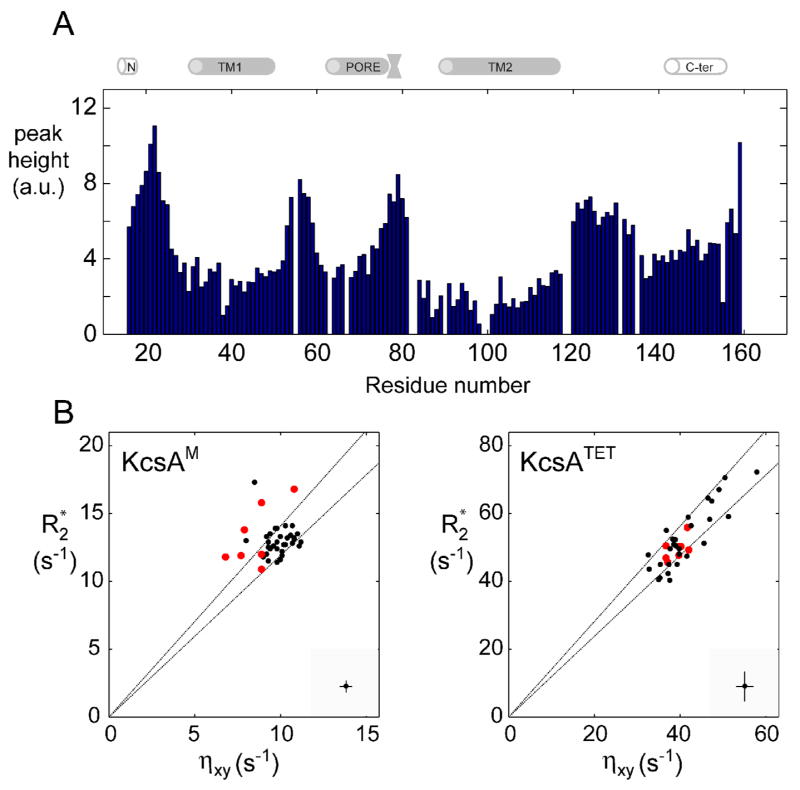
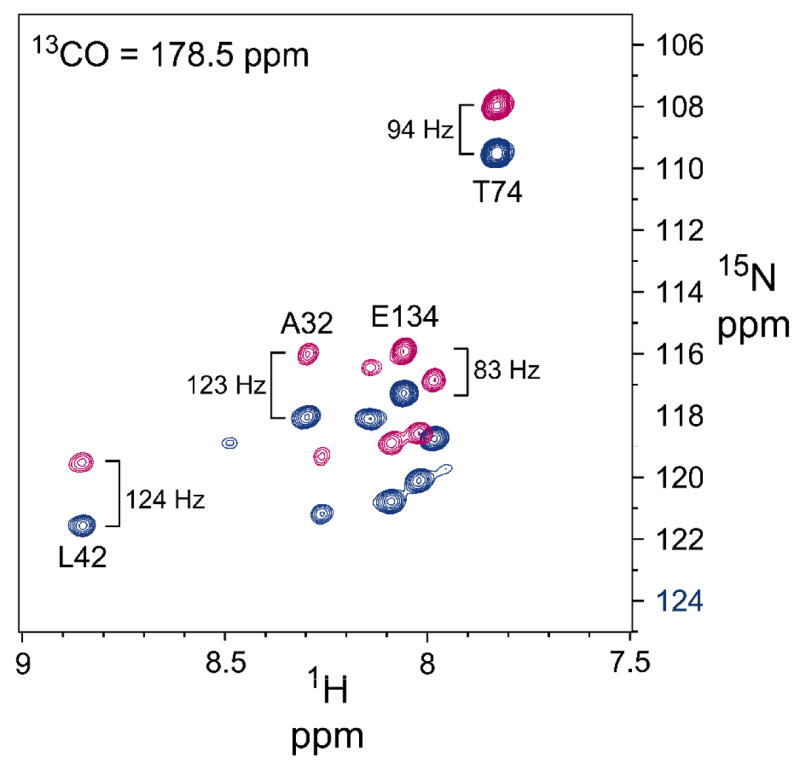
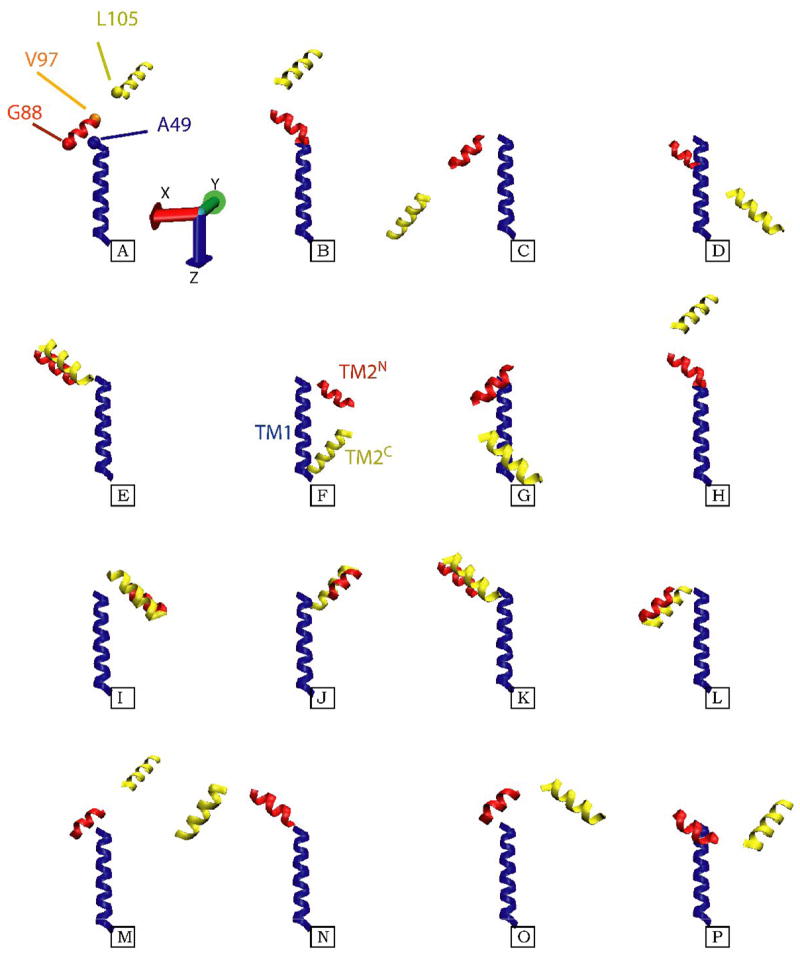
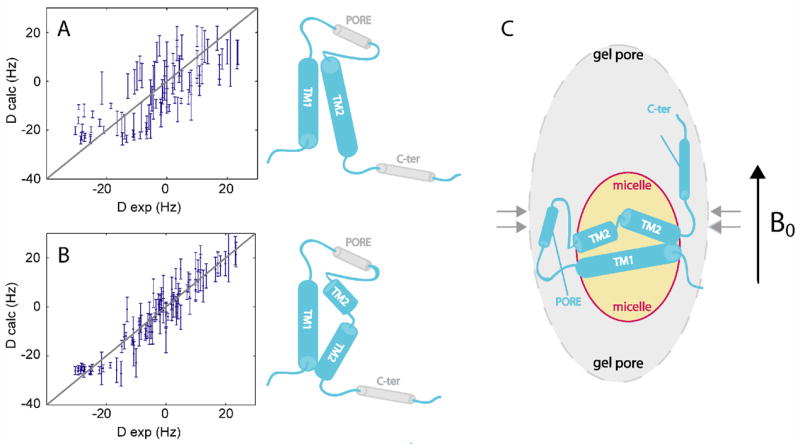
KcsA is a homotetrameric 68-kDa membrane-associated potassium channel which selectively gates the flux of potassium ions across the membrane. The channel is known to undergo a pH-dependent open-to-closed transition. Here we describe an NMR study of the monomeric subunit of the channel (KcsAM), solubilized in SDS micelles. Chemical shift, solvent exchange, backbone 15N relaxation and residual dipolar coupling (RDC) data show the TM1 helix to remain intact, but the TM2 helix contains a distinct kink, which is subject to concentration-independent but pH-dependent conformational exchange on a microsecond time scale. The kink region, centered at G99, was previously implicated in the gating of the tetrameric KcsA channel. An RDC-based model of KcsAM at acidic pH orients TM1 and the two helical segments of the kinked TM2 in a configuration reminiscent of the open conformation of the channel. Thus, the transition between states appears to be an inherent capability of the monomer, with the tetrameric assembly exerting a modulatory effect upon the transition which gives the channel its physiological gating profile.
Figures
Similar articles
-
pH-Dependent Conformational Changes of KcsA Tetramer and Monomer Probed by Raman Spectroscopy.Int J Mol Sci. 2019 Jun 4;20(11):2736. doi: 10.3390/ijms20112736. Int J Mol Sci. 2019. PMID: 31167355 Free PMC article.
-
NMR study of the tetrameric KcsA potassium channel in detergent micelles.Protein Sci. 2006 Apr;15(4):684-98. doi: 10.1110/ps.051954706. Epub 2006 Mar 7. Protein Sci. 2006. PMID: 16522799 Free PMC article.
-
Measurement of 15N relaxation in the detergent-solubilized tetrameric KcsA potassium channel.J Biomol NMR. 2006 Oct;36(2):123-36. doi: 10.1007/s10858-006-9071-4. Epub 2006 Sep 20. J Biomol NMR. 2006. PMID: 17013683
-
The potassium channel KcsA and its interaction with the lipid bilayer.Cell Mol Life Sci. 2003 Aug;60(8):1581-90. doi: 10.1007/s00018-003-3172-y. Cell Mol Life Sci. 2003. PMID: 14513833 Free PMC article. Review.
-
Emerging issues of connexin channels: biophysics fills the gap.Q Rev Biophys. 2001 Aug;34(3):325-472. doi: 10.1017/s0033583501003705. Q Rev Biophys. 2001. PMID: 11838236 Review.
Cited by
-
Conformational heterogeneity in closed and open states of the KcsA potassium channel in lipid bicelles.J Gen Physiol. 2016 Aug;148(2):119-32. doi: 10.1085/jgp.201611602. Epub 2016 Jul 18. J Gen Physiol. 2016. PMID: 27432996 Free PMC article.
-
Paramagnetic-based NMR restraints lift residual dipolar coupling degeneracy in multidomain detergent-solubilized membrane proteins.J Am Chem Soc. 2011 Feb 23;133(7):2232-41. doi: 10.1021/ja109080t. Epub 2011 Feb 2. J Am Chem Soc. 2011. PMID: 21287984 Free PMC article.
-
Recent Advances in the Application of Solution NMR Spectroscopy to Multi-Span Integral Membrane Proteins.Prog Nucl Magn Reson Spectrosc. 2009 Nov 1;55(4):335-360. doi: 10.1016/j.pnmrs.2009.07.002. Prog Nucl Magn Reson Spectrosc. 2009. PMID: 20161395 Free PMC article. No abstract available.
-
Solid-state NMR of the Yersinia pestis outer membrane protein Ail in lipid bilayer nanodiscs sedimented by ultracentrifugation.J Biomol NMR. 2015 Apr;61(3-4):275-86. doi: 10.1007/s10858-014-9893-4. Epub 2015 Jan 13. J Biomol NMR. 2015. PMID: 25578899 Free PMC article.
-
pH-Dependent Conformational Changes of KcsA Tetramer and Monomer Probed by Raman Spectroscopy.Int J Mol Sci. 2019 Jun 4;20(11):2736. doi: 10.3390/ijms20112736. Int J Mol Sci. 2019. PMID: 31167355 Free PMC article.
References
-
- Heginbotham L, Odessey E, Miller C. Tetrameric stoichiometry of a prokaryotic K+ channel. Biochemistry. 1997;36:10335–10342. - PubMed
-
- Doyle DA, Cabral JM, Pfuetzner RA, Kuo A, Gulbis JM, Cohen SL, Chait BT, Mackinnon R. The structure of the potassium channel: molecular basis of K+ conduction and selectivity. Science. 1998;280:69–77. - PubMed
-
- Lange A, Giller K, Hornig S, Martin-Eauclaire MF, Pongs O, Becker S, Baldus M. Toxin-induced conformational changes in a potassium channel revealed by solid-state NMR. Nature. 2006;440:959–962. - PubMed
-
- Yu LP, Sun CH, Song DY, Shen JW, Xu N, Gunasekera A, Hajduk PJ, Olejniczak ET. Nuclear magnetic resonance studies of a potassium channel-charybdotoxin complex. Biochemistry. 2005;44:15834–15841. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
