

Evolution of fitnesses and allele frequencies in a population with spatially heterogeneous selection pressures
- PMID: 17947414
- PMCID: PMC2147970
- DOI: 10.1534/genetics.107.079558
Evolution of fitnesses and allele frequencies in a population with spatially heterogeneous selection pressures
Abstract
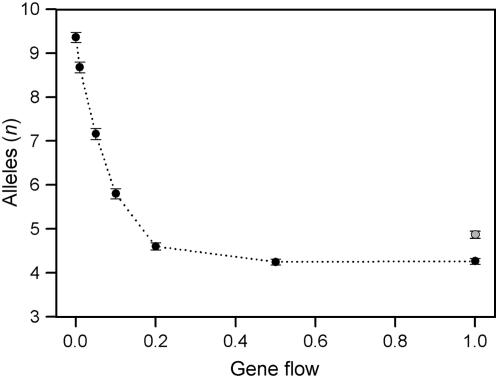
The level of gene flow considerably influences the outcome of evolutionary processes in structured populations with spatial heterogeneity in selection pressures; low levels of gene flow may allow local adaptation whereas high levels of gene flow may oppose this process thus preventing the stable maintenance of polymorphism. Indeed, proportions of fitness space that successfully maintain polymorphism are substantially larger in spatially heterogenous populations with lower to moderate levels of gene flow when compared to single-deme models. Nevertheless, the effect of spatial heterogeneity on the evolutionary construction of polymorphism is less clear. We have investigated the levels of polymorphism resulting from a simple two-deme construction model, which incorporates recurrent mutation as well as selection. We further compared fitness properties, stability of equilibria, and frequency distribution patterns emerging from the construction approach and compared these to the static fitness-space approach. The construction model either promotes or constrains the level of polymorphisms, depending on the levels of gene flow. Comparison of the fitness properties resulting from both approaches shows that they maintain variation in different parts of fitness space. The part of fitness space resulting from construction is more stable than that implied by the ahistoric fitness-space approach. Finally, the equilibrium allele-frequency distribution patterns vary substantially with different levels of gene flow, underlining the importance of correctly sampling spatial structure if these patterns are to be used to estimate population-genetic processes.
Figures






References
-
- Bulmer, M. G., 1972. Multiple niche polymorphism. Am. Nat. 106: 254–257.
-
- Ewens, W. J., 1972. Sampling theory of selectively neutral alleles. Theor. Popul. Biol. 3: 87–112. - PubMed
-
- Felsenstein, J., 1976. The theoretical population genetics of variable selection and migration. Annu. Rev. Genet. 10: 253–280. - PubMed
-
- Gillespie, J. H., 1991. The Causes of Molecular Evolution. Oxford University Press, New York.
-
- Gillespie, J. H., 1997. Junk ain't what junk does: neutral alleles in a selected context. Gene 205: 291–299. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
