

On recombination-induced multiple and simultaneous coalescent events
- PMID: 17947442
- PMCID: PMC2219491
- DOI: 10.1534/genetics.107.071126
On recombination-induced multiple and simultaneous coalescent events
Abstract
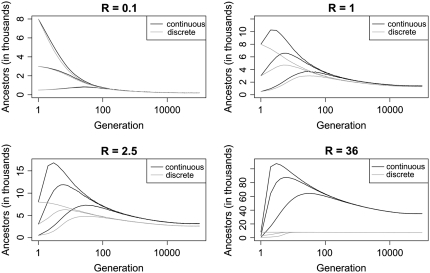
Coalescent theory deals with the dynamics of how sampled genetic material has spread through a population from a single ancestor over many generations and is ubiquitous in contemporary molecular population genetics. Inherent in most applications is a continuous-time approximation that is derived under the assumption that sample size is small relative to the actual population size. In effect, this precludes multiple and simultaneous coalescent events that take place in the history of large samples. If sequences do not recombine, the number of sequences ancestral to a large sample is reduced sufficiently after relatively few generations such that use of the continuous-time approximation is justified. However, in tracing the history of large chromosomal segments, a large recombination rate per generation will consistently maintain a large number of ancestors. This can create a major disparity between discrete-time and continuous-time models and we analyze its importance, illustrated with model parameters typical of the human genome. The presence of gene conversion exacerbates the disparity and could seriously undermine applications of coalescent theory to complete genomes. However, we show that multiple and simultaneous coalescent events influence global quantities, such as total number of ancestors, but have negligible effect on local quantities, such as linkage disequilibrium. Reassuringly, most applications of the coalescent model with recombination (including association mapping) focus on local quantities.
Figures






References
-
- Fu, Y. X., 2006. Exact coalescence for the Wright-Fisher model. Theor. Popul. Biol. 69: 385–394. - PubMed
-
- Griffiths, R. C., and P. Marjoram, 1996. Ancestral inference from samples of DNA sequences with recombination. J. Comput. Biol. 3(4): 479–502. - PubMed
-
- Hein, J., M. H. Schierup and C. Wiuf, 2005. Gene Genealogies Variation and Evolution. Oxford University Press, London/New York/Oxford.
-
- Hudson, R., 1983. Properties of the neutral allele model with intragenic recombination. Theor. Popul. Biol. 23: 183–201. - PubMed
-
- Kingman, J. F. C., 1982. The coalescent. Stoch. Proc. Appl. 13: 235–248.
MeSH terms
LinkOut - more resources
Full Text Sources
