

Histone modifications influence the action of Snf2 family remodelling enzymes by different mechanisms
- PMID: 17949749
- PMCID: PMC2279226
- DOI: 10.1016/j.jmb.2007.09.059
Histone modifications influence the action of Snf2 family remodelling enzymes by different mechanisms
Abstract
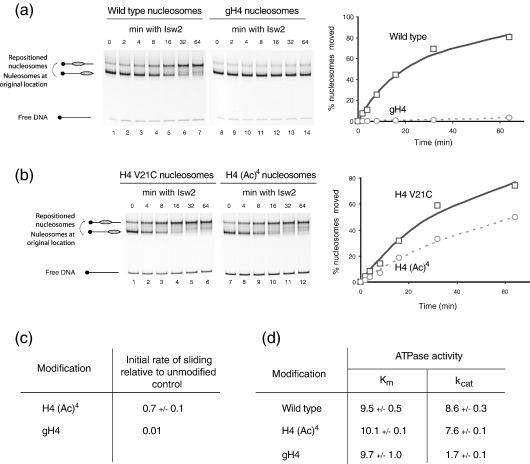
Alteration of chromatin structure by chromatin modifying and remodelling activities is a key stage in the regulation of many nuclear processes. These activities are frequently interlinked, and many chromatin remodelling enzymes contain motifs that recognise modified histones. Here we adopt a peptide ligation strategy to generate specifically modified chromatin templates and used these to study the interaction of the Chd1, Isw2 and RSC remodelling complexes with differentially acetylated nucleosomes. Specific patterns of histone acetylation are found to alter the rate of chromatin remodelling in different ways. For example, histone H3 lysine 14 acetylation acts to increase recruitment of the RSC complex to nucleosomes. However, histone H4 tetra-acetylation alters the spectrum of remodelled products generated by increasing octamer transfer in trans. In contrast, histone H4 tetra-acetylation was also found to reduce the activity of the Chd1 and Isw2 remodelling enzymes by reducing catalytic turnover without affecting recruitment. These observations illustrate a range of different means by which modifications to histones can influence the action of remodelling enzymes.
Figures






Similar articles
-
Histone H3 tail acetylation modulates ATP-dependent remodeling through multiple mechanisms.Nucleic Acids Res. 2011 Oct;39(19):8378-91. doi: 10.1093/nar/gkr535. Epub 2011 Jul 11. Nucleic Acids Res. 2011. PMID: 21749977 Free PMC article.
-
Selective recognition of acetylated histones by bromodomains in transcriptional co-activators.Biochem J. 2007 Feb 15;402(1):125-33. doi: 10.1042/BJ20060907. Biochem J. 2007. PMID: 17049045 Free PMC article.
-
Histone Acetylation near the Nucleosome Dyad Axis Enhances Nucleosome Disassembly by RSC and SWI/SNF.Mol Cell Biol. 2015 Dec;35(23):4083-92. doi: 10.1128/MCB.00441-15. Epub 2015 Sep 28. Mol Cell Biol. 2015. PMID: 26416878 Free PMC article.
-
Snf2-family proteins: chromatin remodellers for any occasion.Curr Opin Chem Biol. 2011 Oct;15(5):649-56. doi: 10.1016/j.cbpa.2011.07.022. Epub 2011 Aug 20. Curr Opin Chem Biol. 2011. PMID: 21862382 Free PMC article. Review.
-
Mechanisms for ATP-dependent chromatin remodelling: the means to the end.FEBS J. 2011 Oct;278(19):3579-95. doi: 10.1111/j.1742-4658.2011.08281.x. Epub 2011 Sep 8. FEBS J. 2011. PMID: 21810178 Free PMC article. Review.
Cited by
-
Distinct amino acids of histone H3 control secondary metabolism in Aspergillus nidulans.Appl Environ Microbiol. 2013 Oct;79(19):6102-9. doi: 10.1128/AEM.01578-13. Epub 2013 Jul 26. Appl Environ Microbiol. 2013. PMID: 23892751 Free PMC article.
-
A role for Snf2-related nucleosome-spacing enzymes in genome-wide nucleosome organization.Science. 2011 Sep 23;333(6050):1758-60. doi: 10.1126/science.1206097. Science. 2011. PMID: 21940898 Free PMC article.
-
Chemical and biological tools for the preparation of modified histone proteins.Top Curr Chem. 2015;363:193-226. doi: 10.1007/128_2015_629. Top Curr Chem. 2015. PMID: 25863817 Free PMC article.
-
Identification and characterization of jasmonic acid- and linolenic acid-mediated transcriptional regulation of secondary laticifer differentiation in Hevea brasiliensis.Sci Rep. 2019 Oct 4;9(1):14296. doi: 10.1038/s41598-019-50800-1. Sci Rep. 2019. PMID: 31586098 Free PMC article.
-
Nucleosomes can invade DNA territories occupied by their neighbors.Nat Struct Mol Biol. 2009 Feb;16(2):151-8. doi: 10.1038/nsmb.1551. Epub 2009 Feb 1. Nat Struct Mol Biol. 2009. PMID: 19182801 Free PMC article.
References
-
- Durr H., Korner C., Muller M., Hickmann V., Hopfner K.P. X-ray structures of the Sulfolobus solfataricus SWI2/SNF2 ATPase core and its complex with DNA. Cell. 2005;121:363–373. - PubMed
-
- Thoma N.H., Czyzewski B.K., Alexeev A.A., Mazin A.V., Kowalczykowski S.C., Pavletich N.P. Structure of the SWI2/SNF2 chromatin-remodeling domain of eukaryotic Rad54. Nature Struct. Mol. Biol. 2005;12:350–356. - PubMed
-
- Saha A., Wittmeyer J., Cairns B.R. Chromatin remodelling: the industrial revolution of DNA around histones. Nature Rev. Mol. Cell Biol. 2006;7:437–447. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
