

Identification of two gene clusters and a transcriptional regulator required for Pseudomonas aeruginosa glycine betaine catabolism
- PMID: 17951379
- PMCID: PMC2293255
- DOI: 10.1128/JB.01393-07
Identification of two gene clusters and a transcriptional regulator required for Pseudomonas aeruginosa glycine betaine catabolism
Abstract
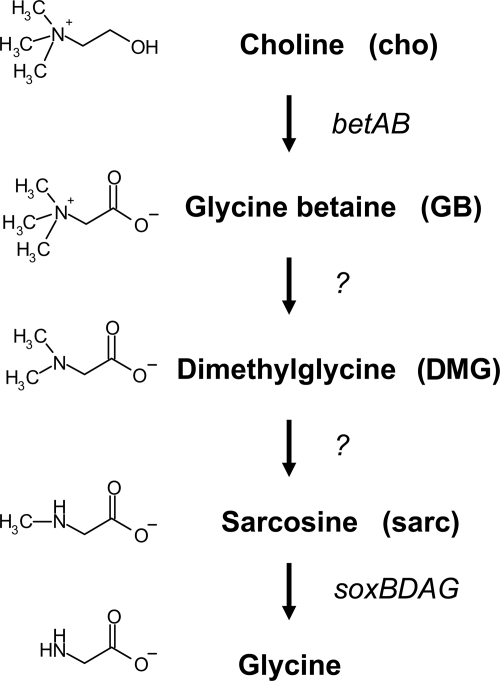
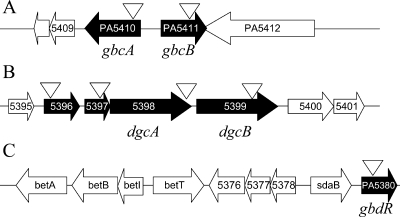
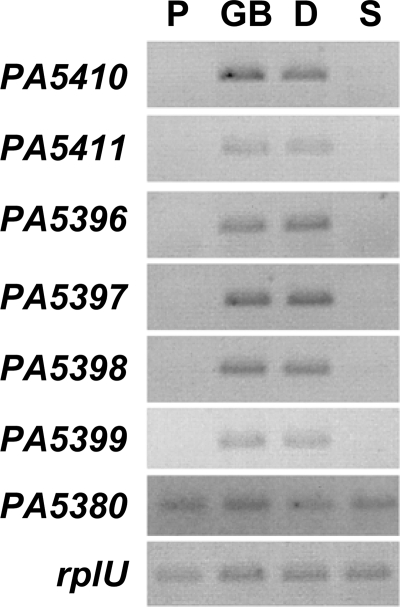
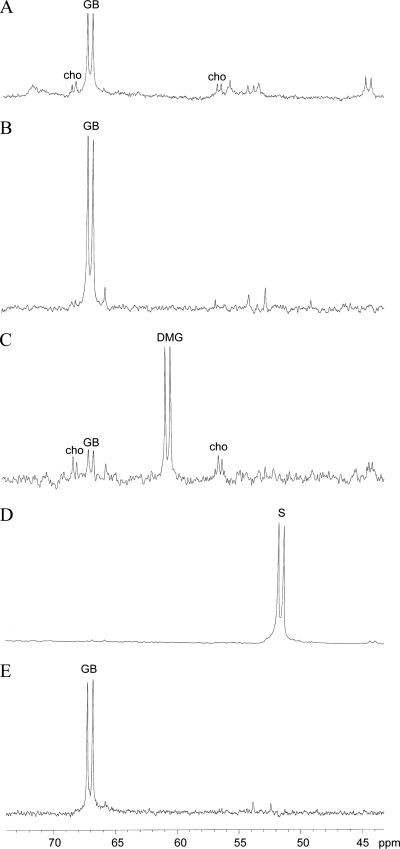
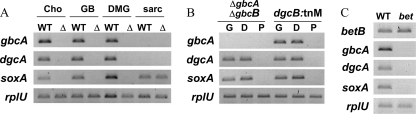
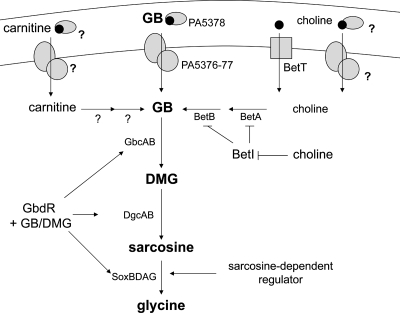
Glycine betaine (GB), which occurs freely in the environment and is an intermediate in the catabolism of choline and carnitine, can serve as a sole source of carbon or nitrogen in Pseudomonas aeruginosa. Twelve mutants defective in growth on GB as the sole carbon source were identified through a genetic screen of a nonredundant PA14 transposon mutant library. Further growth experiments showed that strains with mutations in two genes, gbcA (PA5410) and gbcB (PA5411), were capable of growth on dimethylglycine (DMG), a catabolic product of GB, but not on GB itself. Subsequent nuclear magnetic resonance (NMR) experiments with 1,2-(13)C-labeled choline indicated that these genes are necessary for conversion of GB to DMG. Similar experiments showed that strains with mutations in the dgcAB (PA5398-PA5399) genes, which exhibit homology to genes that encode other enzymes with demethylase activity, are required for the conversion of DMG to sarcosine. Mutant analyses and (13)C NMR studies also confirmed that the soxBDAG genes, predicted to encode a sarcosine oxidase, are required for sarcosine catabolism. Our screen also identified a predicted AraC family transcriptional regulator, encoded by gbdR (PA5380), that is required for growth on GB and DMG and for the induction of gbcA, gbcB, and dgcAB in response to GB or DMG. Mutants defective in the previously described gbt gene (PA3082) grew on GB with kinetics similar to those of the wild type in both the PAO1 and PA14 strain backgrounds. These studies provided important insight into both the mechanism and the regulation of the catabolism of GB in P. aeruginosa.
Figures
References
-
- Bazire, A., A. Dheilly, F. Diab, D. Morin, M. Jebbar, D. Haras, and A. Dufour. 2005. Osmotic stress and phosphate limitation alter production of cell-to-cell signal molecules and rhamnolipid biosurfactant by Pseudomonas aeruginosa. FEMS Microbiol. Lett. 253125-131. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
