

Establishment of the ambient pH signaling complex in Aspergillus nidulans: PalI assists plasma membrane localization of PalH
- PMID: 17951518
- PMCID: PMC2168248
- DOI: 10.1128/EC.00275-07
Establishment of the ambient pH signaling complex in Aspergillus nidulans: PalI assists plasma membrane localization of PalH
Abstract
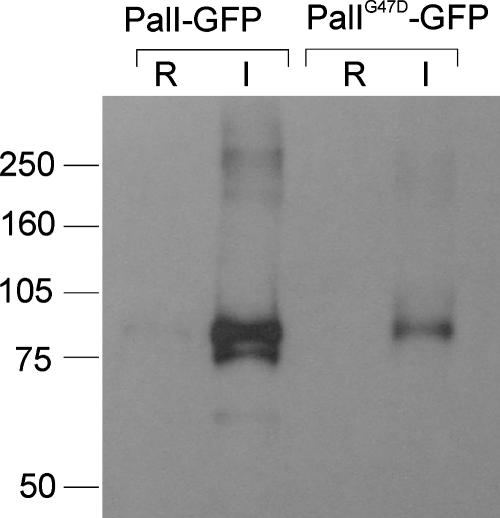
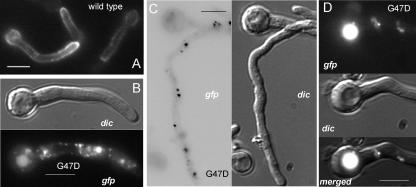
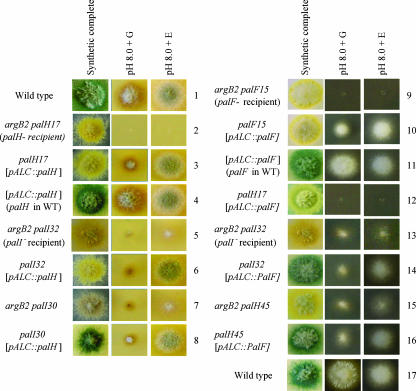
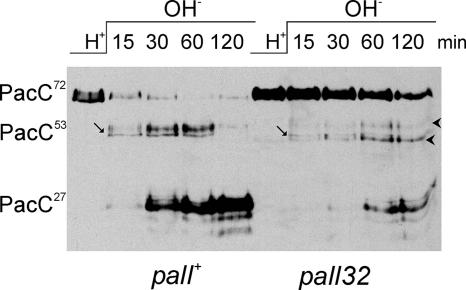
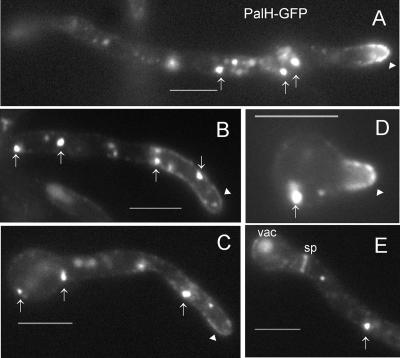
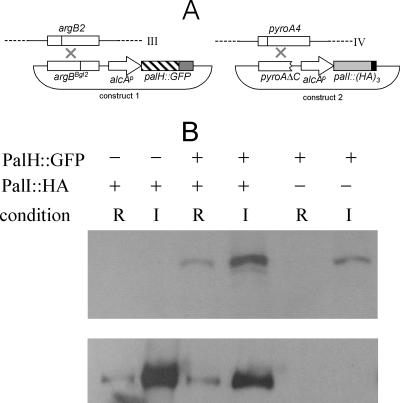
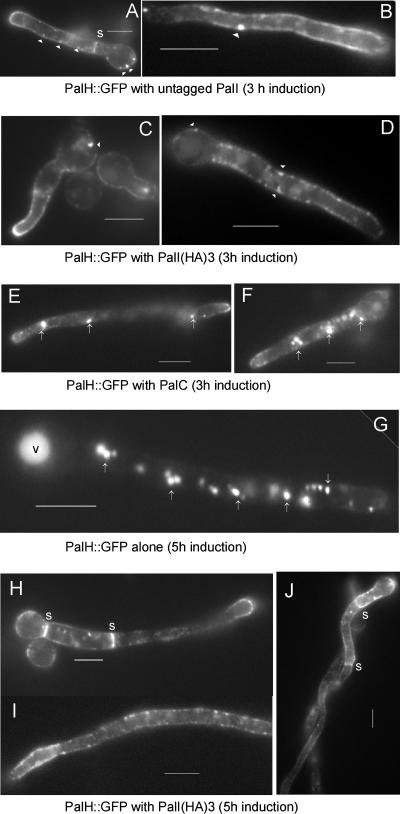
The Aspergillus nidulans ambient pH signaling pathway involves two transmembrane domain (TMD)-containing proteins, PalH and PalI. We provide in silico and mutational evidence suggesting that PalI is a three TMD (3-TMD) protein with an N-terminal signal peptide, and we show that PalI localizes to the plasma membrane. PalI is not essential for the proteolytic conversion of the PacC translation product into the processed 27-kDa form, but its absence markedly reduces the accumulation of the 53-kDa intermediate after cells are shifted to an alkaline pH. PalI and its homologues contain a predicted luminal, conserved Gly-Cys-containing motif that distantly resembles a Gly-rich dimerization domain. The Gly44Arg and Gly47Asp substitutions within this motif lead to loss of function. The Gly47Asp substitution prevents plasma membrane localization of PalI-green fluorescent protein (GFP) and leads to its missorting into the multivesicular body pathway. Overexpression of the likely ambient alkaline pH receptor, the 7-TMD protein PalH, partially suppresses the null palI32 mutation. Although some PalH-GFP localizes to the plasma membrane, it predominates in internal membranes. However, the coexpression of PalI to stoichiometrically similar levels results in the strong predominance of PalH-GFP in the plasma membrane. Thus, one role for PalI, but possibly not the only role, is to assist with plasma membrane localization of PalH. These data, considered along with previous reports for both Saccharomyces cerevisiae and A. nidulans, strongly support the prevailing model of pH signaling involving two spatially segregated complexes: a plasma membrane complex containing PalH, PalI, and the arrestin-like protein PalF and an endosomal membrane complex containing PalA and PalB, to which PacC is recruited for its proteolytic activation.
Figures

References
-
- Arst, H. N., Jr., E. Bignell, and J. Tilburn. 1994. Two new genes involved in signaling ambient pH in Aspergillus nidulans. Mol. Gen. Genet. 245:787-790. - PubMed
-
- Arst, H. N., Jr., and M. A. Peñalva. 2003. pH regulation in Aspergillus and parallels with higher eukaryotic regulatory systems. Trends Genet. 19:224-231. - PubMed
-
- Babst, M., D. J. Katzmann, E. J. Estepa-Sabal, T. Meerloo, and S. D. Emr. 2002. ESCRT-III: an endosome-associated heterooligomeric protein complex required for MVB sorting. Dev. Cell 3:271-282. - PubMed
-
- Bartnicki-García, S., F. Hergert, and G. Gierz. 1989. Computer simulation of fungal morphogenesis and the mathematical basis for hyphal tip growth. Protoplasma 153:46-57.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
