

An overview of structural DNA nanotechnology
- PMID: 17952671
- PMCID: PMC3479651
- DOI: 10.1007/s12033-007-0059-4
An overview of structural DNA nanotechnology
Abstract
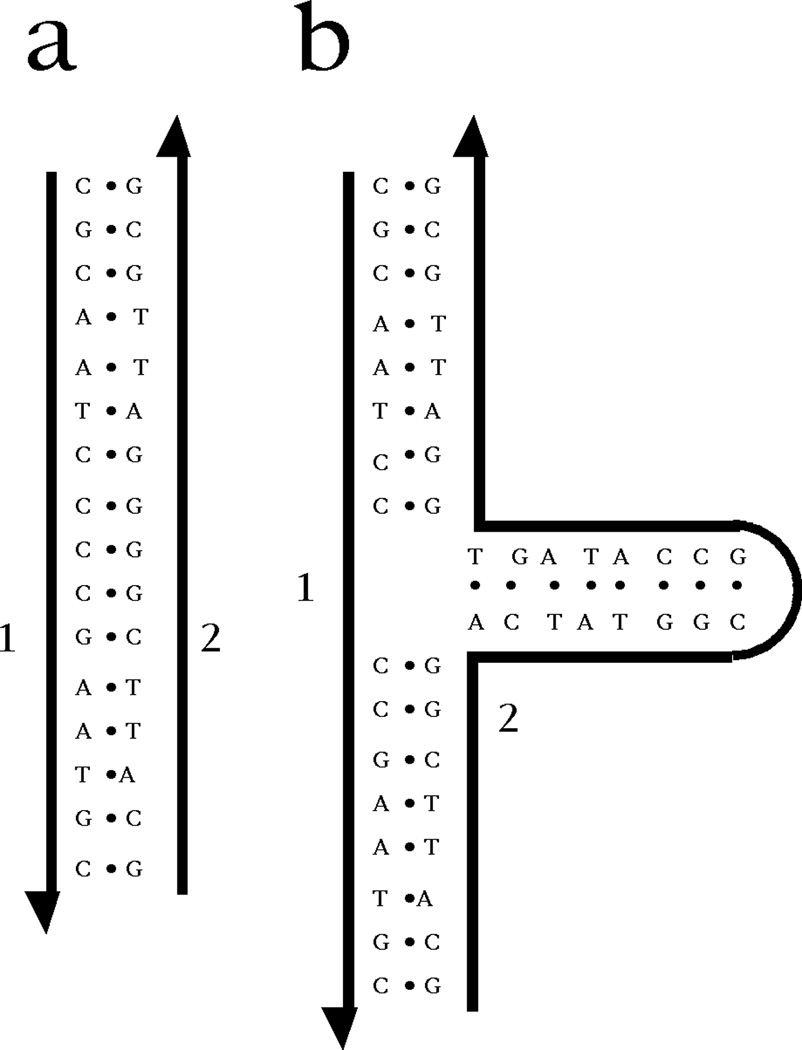
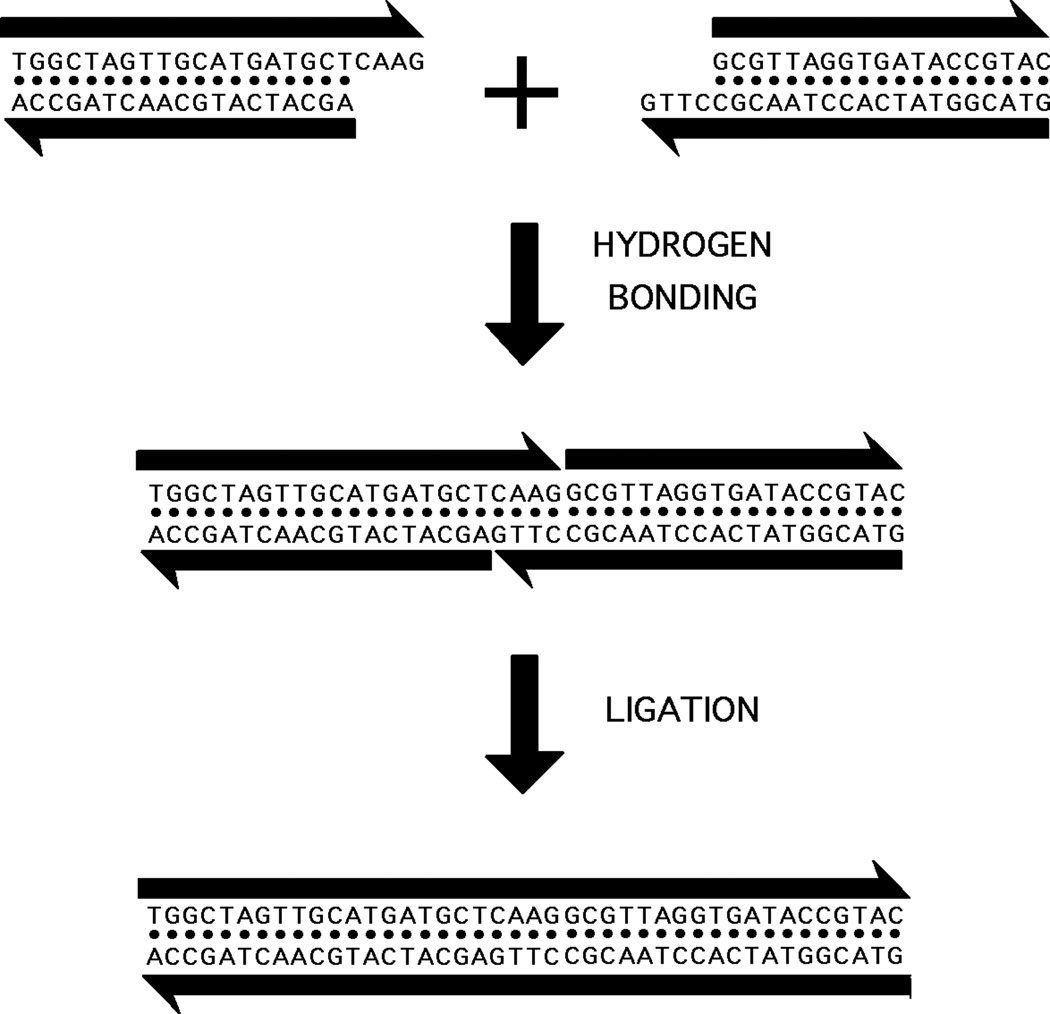
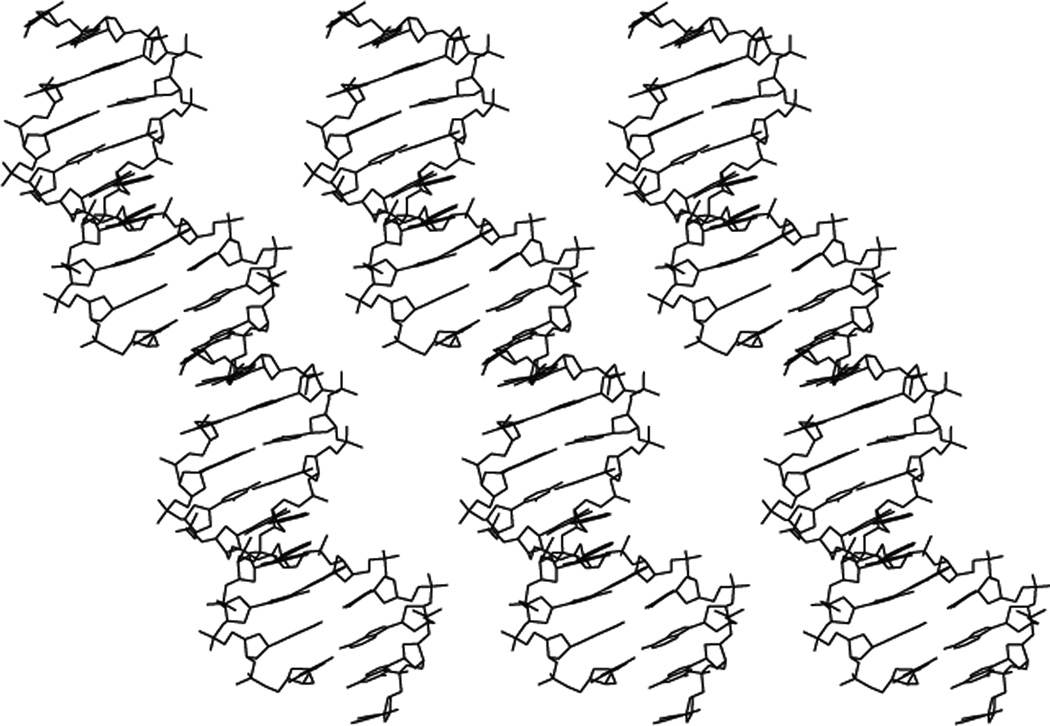
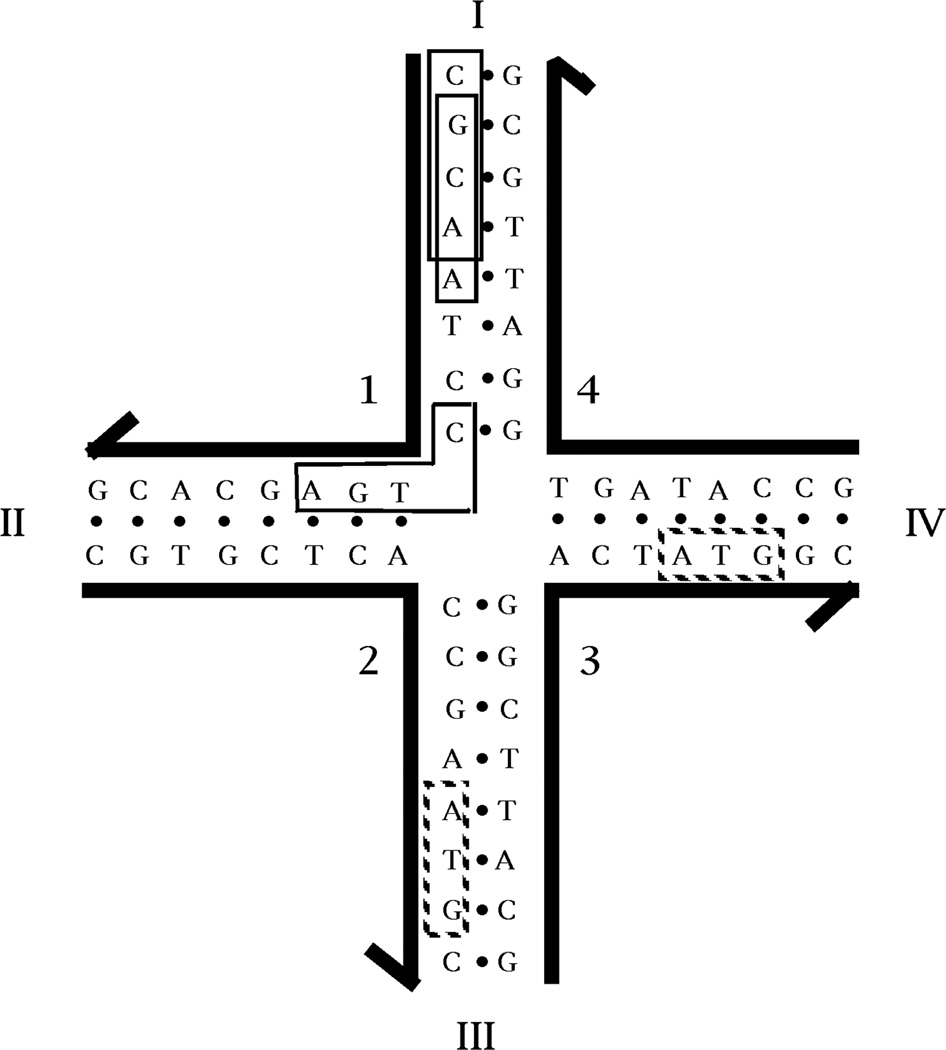
Structural DNA Nanotechnology uses unusual DNA motifs to build target shapes and arrangements. These unusual motifs are generated by reciprocal exchange of DNA backbones, leading to branched systems with many strands and multiple helical domains. The motifs may be combined by sticky ended cohesion, involving hydrogen bonding or covalent interactions. Other forms of cohesion involve edge-sharing or paranemic interactions of double helices. A large number of individual species have been developed by this approach, including polyhedral catenanes, a variety of single-stranded knots, and Borromean rings. In addition to these static species, DNA-based nanomechanical devices have been produced that are ultimately targeted to lead to nanorobotics. Many of the key goals of structural DNA nanotechnology entail the use of periodic arrays. A variety of 2D DNA arrays have been produced with tunable features, such as patterns and cavities. DNA molecules have be used successfully in DNA-based computation as molecular representations of Wang tiles, whose self-assembly can be programmed to perform a calculation. About 4 years ago, on the fiftieth anniversary of the double helix, the area appeared to be at the cusp of a truly exciting explosion of applications; this was a correct assessment, and much progress has been made in the intervening period.
Figures







References
-
- Watson JD, Crick FHC. A structure for deoxyribose nucleic acid. Nature. 1953;171:737–738. - PubMed
-
- Seeman NC. Nucleic acid junctions and lattices. J. Theor. Biol. 1982;99:237–247. - PubMed
-
- Robinson BH, Seeman NC. The design of a biochip. Prot. Eng. 1987;1:295–300. - PubMed
-
- Winfree E. On the computational power of DNA annealing and ligation. In: Lipton EJ, Baum EB, editors. DNA Based Computing. 1996. pp. 199–219. Am. Math. Soc., Providence.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
