

Cordon-bleu is an actin nucleation factor and controls neuronal morphology
- PMID: 17956734
- PMCID: PMC2507594
- DOI: 10.1016/j.cell.2007.08.030
Cordon-bleu is an actin nucleation factor and controls neuronal morphology
Abstract
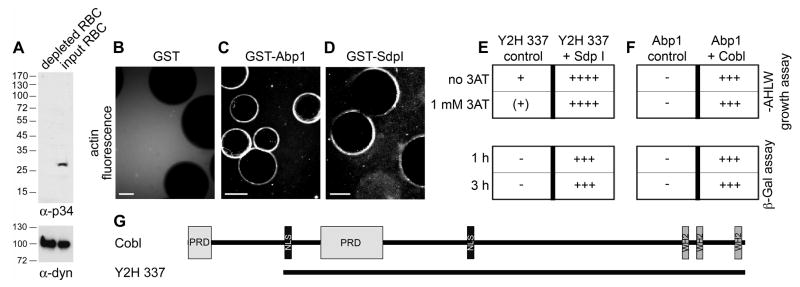
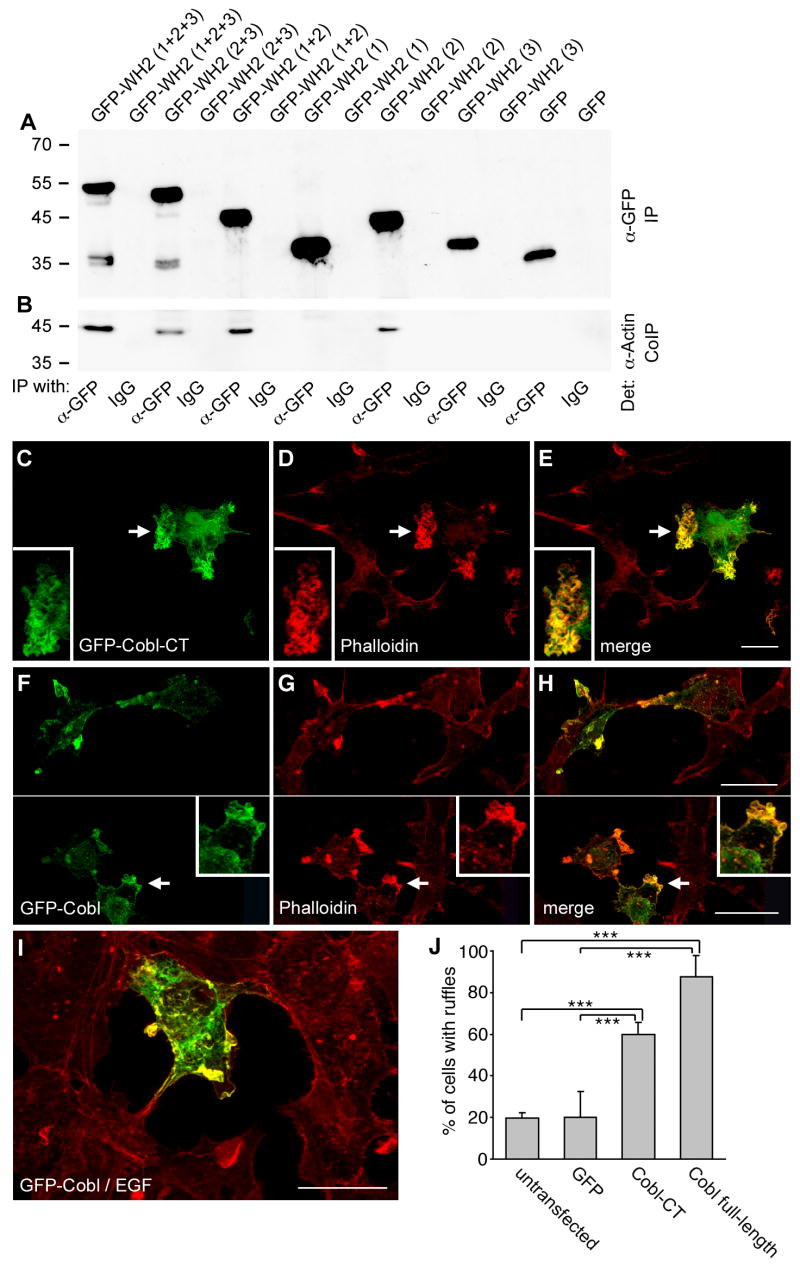
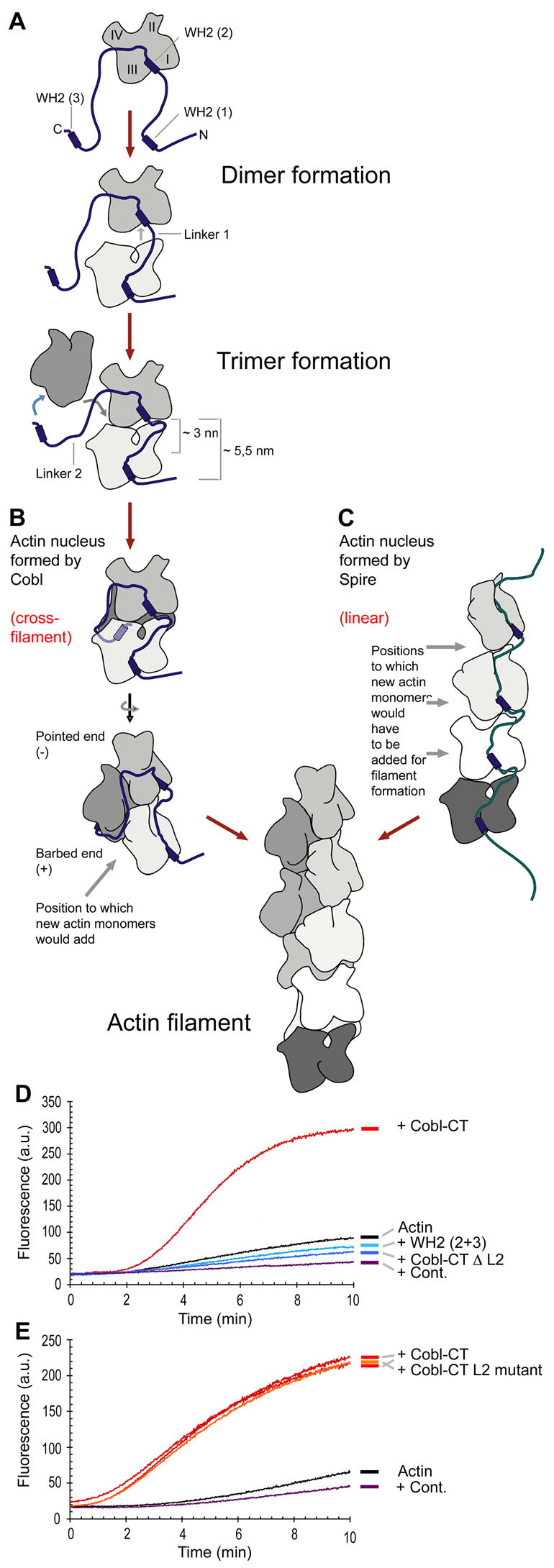
Despite the wealth of different actin structures formed, only two actin nucleation factors are well established in vertebrates: the Arp2/3 complex and formins. Here, we describe a further nucleator, cordon-bleu (Cobl). Cobl is a brain-enriched protein using three Wiskott-Aldrich syndrome protein homology 2 (WH2) domains for actin binding. Cobl promotes nonbundled, unbranched filaments. Filament formation relies on barbed-end growth and requires all three Cobl WH2 domains and the extended linker L2. We suggest that the nucleation power of Cobl is based on the assembly of three actin monomers in cross-filament orientation. Cobl localizes to sites of high actin dynamics and modulates cell morphology. In neurons, induction of both neurites and neurite branching is dramatically increased by Cobl expression-effects that critically depend on Cobl's actin nucleation ability. Correspondingly, Cobl depletion results in decreased dendritic arborization. Thus, Cobl is an actin nucleator controlling neuronal morphology and development.
Figures




Comment in
-
Cordon-bleu: a new taste in actin nucleation.Cell. 2007 Oct 19;131(2):236-8. doi: 10.1016/j.cell.2007.10.003. Cell. 2007. PMID: 17956726
References
-
- Blanchoin L, Amann KJ, Higgs HN, Marchand JB, Kaiser DA, Pollard TD. Direct observation of dendritic actin filament networks nucleated by Arp2/3 complex and WASP/Scar proteins. Nature. 2000;404:1007–11. - PubMed
-
- Carroll EA, Gerrelli D, Gasca S, Berg E, Beier DR, Copp AJ, Klingensmith J. Cordon-bleu is a conserved gene involved in neural tube formation. Dev Biol. 2003;262:16–31. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
