

Gene expression underlying adaptive variation in Heliconius wing patterns: non-modular regulation of overlapping cinnabar and vermilion prepatterns
- PMID: 17956848
- PMCID: PMC2562400
- DOI: 10.1098/rspb.2007.1115
Gene expression underlying adaptive variation in Heliconius wing patterns: non-modular regulation of overlapping cinnabar and vermilion prepatterns
Abstract
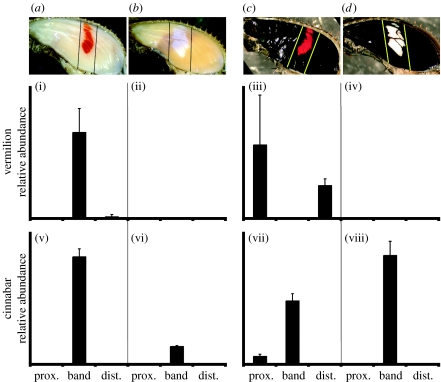
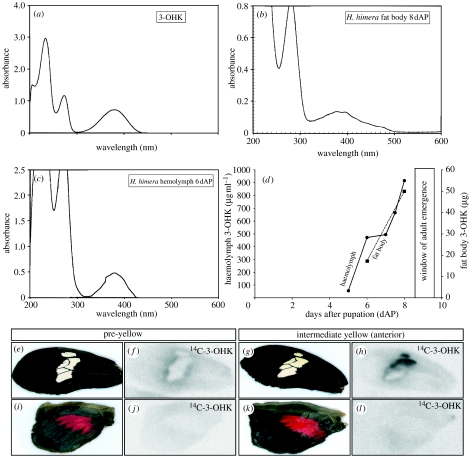
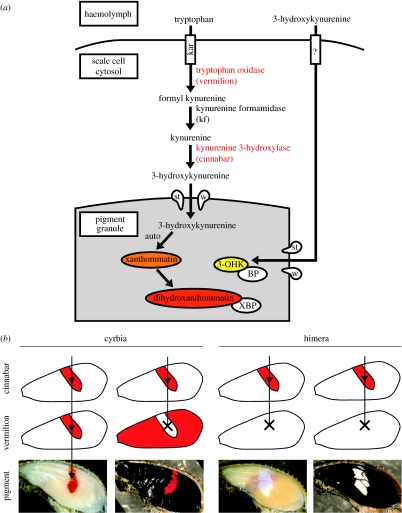
Geographical variation in the mimetic wing patterns of the butterfly Heliconius erato is a textbook example of adaptive polymorphism; however, little is known about how this variation is controlled developmentally. Using microarrays and qPCR, we identified and compared expression of candidate genes potentially involved with a red/yellow forewing band polymorphism in H. erato. We found that transcripts encoding the pigment synthesis enzymes cinnabar and vermilion showed pattern- and polymorphism-related expression patterns, respectively. cinnabar expression was associated with the forewing band regardless of pigment colour, providing the first gene expression pattern known to be correlated with a major Heliconius colour pattern. In contrast, vermilion expression changed spatially over time in red-banded butterflies, but was not expressed at detectable levels in yellow-banded butterflies, suggesting that regulation of this gene may be involved with the red/yellow polymorphism. Furthermore, we found that the yellow pigment, 3-hydroxykynurenine, is incorporated into wing scales from the haemolymph rather than being synthesized in situ. We propose that some aspects of Heliconius colour patterns are determined by spatio-temporal overlap of pigment gene transcription prepatterns and speculate that evolutionary changes in vermilion regulation may in part underlie an adaptive colour pattern polymorphism.
Figures

References
-
- Benson W.W. Natural selection for Müllerian mimicry in Heliconius erato in Costa Rica. Science. 1972;176:936–939. doi:10.1126/science.176.4037.936 - DOI - PubMed
-
- Brown, K. S. J. 1979 Ecologia Geográfica e Evolução nas Florestas Neotropicais PhD dissertation. Universidade Estadual de Campinas, Campinas.
-
- Flanagan N.S, Tobler A, Davison A, Pybus O.G, Kapan D.D, Planas S, Linares M, Heckel D, McMillan W.O. Historical demography of Mullerian mimicry in the neotropical Heliconius butterflies. Proc. Natl Acad. Sci. USA. 2004;101:9704–9709. doi:10.1073/pnas.0306243101 - DOI - PMC - PubMed
-
- Gilbert L.E. Adaptive novelty through introgression in Heliconius wing patterns: evidence for a shared genetic “toolbox” from synthetic hybrid zones and a theory of diversification. In: Boogs C.L, Watt W.B, Ehrlich P.R, editors. Butterflies: ecology and evolution taking flight. The University of Chicago Press; Chicago, IL: 2003.
-
- Gilbert L.E, Forrest H.S, Schultz T.D, Harvey D.J. Correlations of ultrastructure and pigmentation suggest how genes control development of wing scale of Heliconius butterflies. J. Res. Lepidopt. 1988;26:141–160.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
