

K13 blocks KSHV lytic replication and deregulates vIL6 and hIL6 expression: a model of lytic replication induced clonal selection in viral oncogenesis
- PMID: 17957251
- PMCID: PMC2020437
- DOI: 10.1371/journal.pone.0001067
K13 blocks KSHV lytic replication and deregulates vIL6 and hIL6 expression: a model of lytic replication induced clonal selection in viral oncogenesis
Abstract
Background: Accumulating evidence suggests that dysregulated expression of lytic genes plays an important role in KSHV (Kaposi's sarcoma associated herpesvirus) tumorigenesis. However, the molecular events leading to the dysregulation of KSHV lytic gene expression program are incompletely understood.
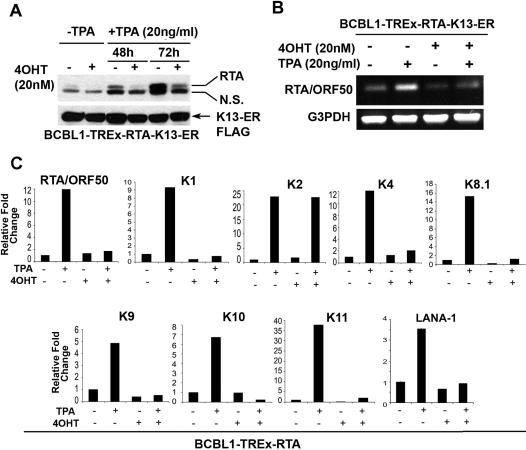
Methodology/principal findings: We have studied the effect of KSHV-encoded latent protein vFLIP K13, a potent activator of the NF-kappaB pathway, on lytic reactivation of the virus. We demonstrate that K13 antagonizes RTA, the KSHV lytic-regulator, and effectively blocks the expression of lytic proteins, production of infectious virions and death of the infected cells. Induction of lytic replication selects for clones with increased K13 expression and NF-kappaB activity, while siRNA-mediated silencing of K13 induces the expression of lytic genes. However, the suppressive effect of K13 on RTA-induced lytic genes is not uniform and it fails to block RTA-induced viral IL6 secretion and cooperates with RTA to enhance cellular IL-6 production, thereby dysregulating the lytic gene expression program.
Conclusions/significance: Our results support a model in which ongoing KSHV lytic replication selects for clones with progressively higher levels of K13 expression and NF-kappaB activity, which in turn drive KSHV tumorigenesis by not only directly stimulating cellular survival and proliferation, but also indirectly by dysregulating the viral lytic gene program and allowing non-lytic production of growth-promoting viral and cellular genes. Lytic Replication-Induced Clonal Selection (LyRICS) may represent a general mechanism in viral oncogenesis.
Conflict of interest statement
Figures







Similar articles
-
Genome-Wide Identification of Direct RTA Targets Reveals Key Host Factors for Kaposi's Sarcoma-Associated Herpesvirus Lytic Reactivation.J Virol. 2019 Feb 19;93(5):e01978-18. doi: 10.1128/JVI.01978-18. Print 2019 Mar 1. J Virol. 2019. PMID: 30541837 Free PMC article.
-
A nuclear role for Kaposi's sarcoma-associated herpesvirus-encoded K13 protein in gene regulation.Oncogene. 2008 Sep 4;27(39):5243-53. doi: 10.1038/onc.2008.150. Epub 2008 May 12. Oncogene. 2008. PMID: 18469854
-
Kaposi's sarcoma-associated herpesvirus-encoded latency-associated nuclear antigen inhibits lytic replication by targeting Rta: a potential mechanism for virus-mediated control of latency.J Virol. 2004 Jun;78(12):6585-94. doi: 10.1128/JVI.78.12.6585-6594.2004. J Virol. 2004. PMID: 15163750 Free PMC article.
-
The regulation of KSHV lytic reactivation by viral and cellular factors.Curr Opin Virol. 2022 Feb;52:39-47. doi: 10.1016/j.coviro.2021.11.004. Epub 2021 Dec 3. Curr Opin Virol. 2022. PMID: 34872030 Free PMC article. Review.
-
Reactivation and Lytic Replication of Kaposi's Sarcoma-Associated Herpesvirus: An Update.Front Microbiol. 2017 Apr 20;8:613. doi: 10.3389/fmicb.2017.00613. eCollection 2017. Front Microbiol. 2017. PMID: 28473805 Free PMC article. Review.
Cited by
-
Deletion of Kaposi's sarcoma-associated herpesvirus FLICE inhibitory protein, vFLIP, from the viral genome compromises the activation of STAT1-responsive cellular genes and spindle cell formation in endothelial cells.J Virol. 2011 Oct;85(19):10375-88. doi: 10.1128/JVI.00226-11. Epub 2011 Jul 27. J Virol. 2011. PMID: 21795355 Free PMC article.
-
Kaposi's sarcoma associated herpesvirus encoded viral FLICE inhibitory protein K13 activates NF-κB pathway independent of TRAF6, TAK1 and LUBAC.PLoS One. 2012;7(5):e36601. doi: 10.1371/journal.pone.0036601. Epub 2012 May 8. PLoS One. 2012. PMID: 22590573 Free PMC article.
-
Targeting Myc in KSHV-associated primary effusion lymphoma with BET bromodomain inhibitors.Oncogene. 2014 May 29;33(22):2928-37. doi: 10.1038/onc.2013.242. Epub 2013 Jun 24. Oncogene. 2014. PMID: 23792448 Free PMC article.
-
Mechanisms of Kaposi's Sarcoma-Associated Herpesvirus Latency and Reactivation.Adv Virol. 2011;2011:193860. doi: 10.1155/2011/193860. Adv Virol. 2011. PMID: 21625290 Free PMC article.
-
Kaposi's sarcoma-associated herpesvirus ORF57 promotes escape of viral and human interleukin-6 from microRNA-mediated suppression.J Virol. 2011 Mar;85(6):2620-30. doi: 10.1128/JVI.02144-10. Epub 2011 Jan 5. J Virol. 2011. PMID: 21209110 Free PMC article.
References
-
- Chang Y, Cesarman E, Pessin MS, Lee F, Culpepper J, et al. Identification of herpesvirus-like DNA sequences in AIDS-associated Kaposi's sarcoma. Science. 1994;266:1865–1869. - PubMed
-
- Cesarman E, Chang Y, Moore PS, Said JW, Knowles DM. Kaposi's sarcoma-associated herpesvirus-like DNA sequences in AIDS- related body-cavity-based lymphomas. N Engl J Med. 1995;332:1186–1191. - PubMed
-
- Soulier J, Grollet L, Oksenhendler E, Cacoub P, Cazals-Hatem D, et al. Kaposi's sarcoma-associated herpesvirus-like DNA sequences in multicentric Castleman's disease. Blood. 1995;86:1276–1280. - PubMed
-
- Nicholas J. Human herpesvirus 8-encoded proteins with potential roles in virus-associated neoplasia. Front Biosci. 2007;12:265–281. - PubMed
-
- Montaner S, Sodhi A, Molinolo A, Bugge TH, Sawai ET, et al. Endothelial infection with KSHV genes in vivo reveals that vGPCR initiates Kaposi's sarcomagenesis and can promote the tumorigenic potential of viral latent genes. Cancer Cell. 2003;3:23–36. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
