

Mitochondrial P450-dependent arachidonic acid metabolism by TCDD-induced hepatic CYP1A5; conversion of EETs to DHETs by mitochondrial soluble epoxide hydrolase
- PMID: 17959137
- PMCID: PMC2868376
- DOI: 10.1016/j.abb.2007.08.012
Mitochondrial P450-dependent arachidonic acid metabolism by TCDD-induced hepatic CYP1A5; conversion of EETs to DHETs by mitochondrial soluble epoxide hydrolase
Abstract
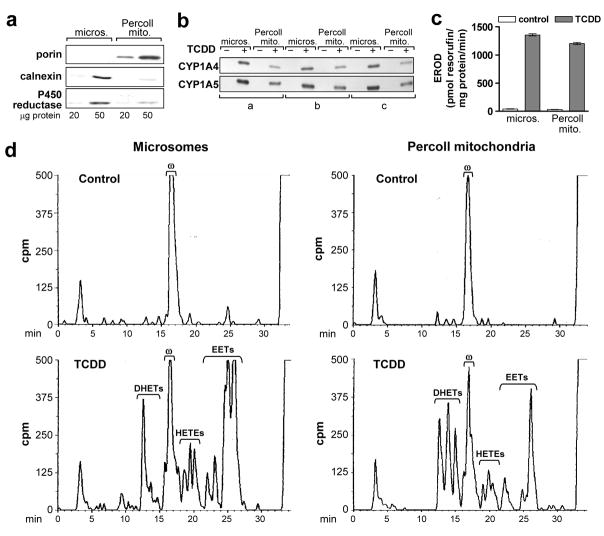
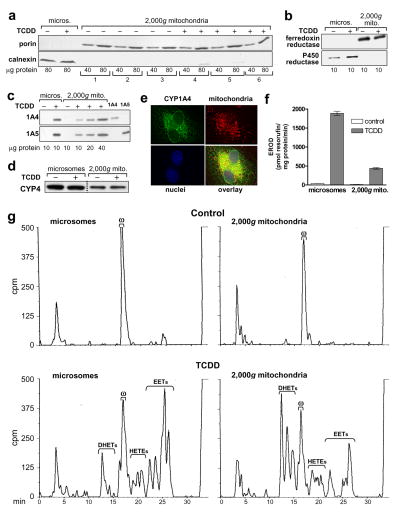
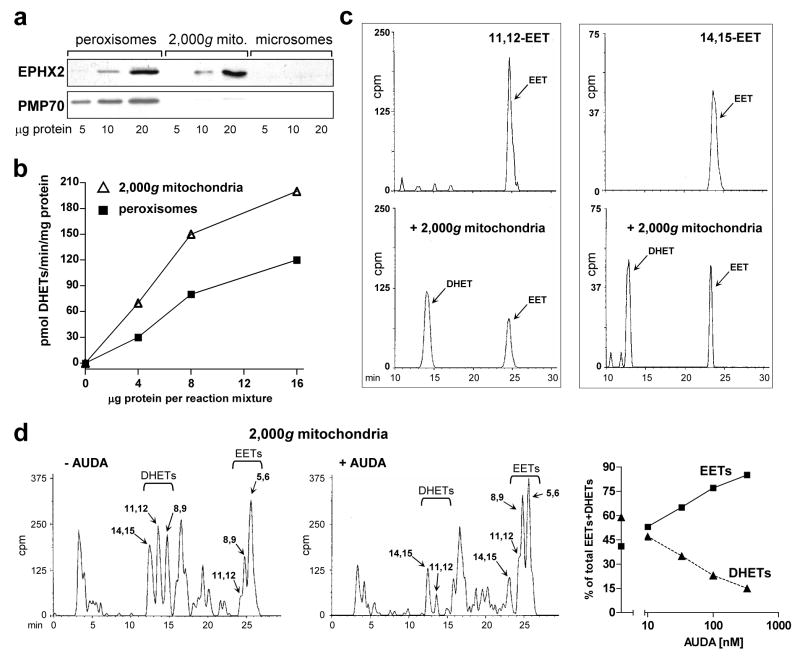
Several P450 enzymes localized in the endoplasmic reticulum and thought to be involved primarily in xenobiotic metabolism, including mouse and rat CYP1A1 and mouse CYP1A2, have also been found to translocate to mitochondria. We report here that the environmental toxin 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) induces enzymatically active CYP1A4/1A5, the avian orthologs of mammalian CYP1A1/1A2, in chick embryo liver mitochondria as well as in microsomes. P450 proteins and activity levels (CYP1A4-dependent 7-ethoxyresorufin-O-deethylase and CYP1A5-dependent arachidonic acid epoxygenation) in mitochondria were 23-40% of those in microsomes. DHET formation by mitochondria was twice that of microsomes and was attributable to a mitochondrial soluble epoxide hydrolase as confirmed by Western blotting with antiEPHX2, conversion by mitochondria of pure 11,12 and 14,15-EET to the corresponding DHETs and inhibition of DHET formation by the soluble epoxide hydrolase inhibitor, 12(-3-adamantan-1-yl-ureido)-dodecanoic acid (AUDA). TCDD also suppressed formation of mitochondrial and microsomal 20-HETE. The findings newly identify mitochondria as a site of P450-dependent arachidonic acid metabolism and as a potential target for TCDD effects. They also demonstrate that mitochondria contain soluble epoxide hydrolase and underscore a role for CYP1A in endobiotic metabolism.
Figures


References
-
- Rifkind AB. CYP1A in TCDD toxicity and in physiology-with particular reference to CYP dependent arachidonic acid metabolism and other endogenous substrates. Drug Metab Rev. 2006;38:291–335. - PubMed
-
- Omura T. Mitochondrial P450s. Chem Biol Interact. 2006;163:86–93. - PubMed
-
- Anandatheerthavarada HK, Vijayasarathy C, Bhagwat SV, Biswas G, Mullick J, Avadhani NG. Physiological role of the N-terminal processed P4501A1 targeted to mitochondria in erythromycin metabolism and reversal of erythromycin-mediated inhibition of mitochondrial protein synthesis. J Biol Chem. 1999;274:6617–6625. - PubMed
-
- Bhagwat SV, Mullick J, Raza H, Avadhani NG. Constitutive and inducible cytochromes P450 in rat lung mitochondria: xenobiotic induction, relative abundance, and catalytic properties. Toxicol Appl Pharmacol. 1999;156:231–240. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
