

Dominance of the proximal coordinate frame in determining the locations of hippocampal place cell activity during navigation
- PMID: 17959742
- PMCID: PMC2268613
- DOI: 10.1152/jn.00731.2007
Dominance of the proximal coordinate frame in determining the locations of hippocampal place cell activity during navigation
Abstract
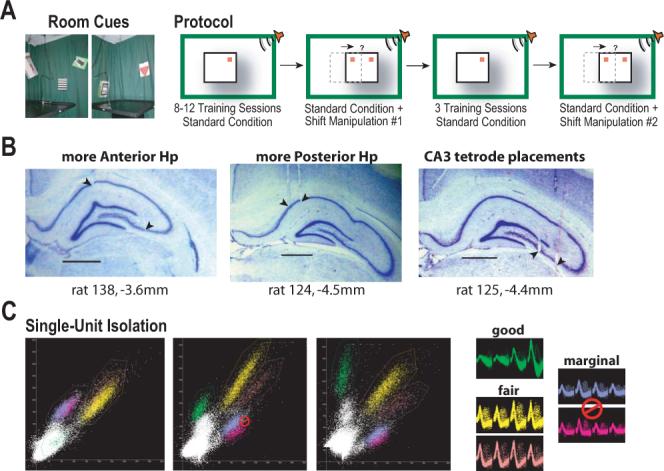
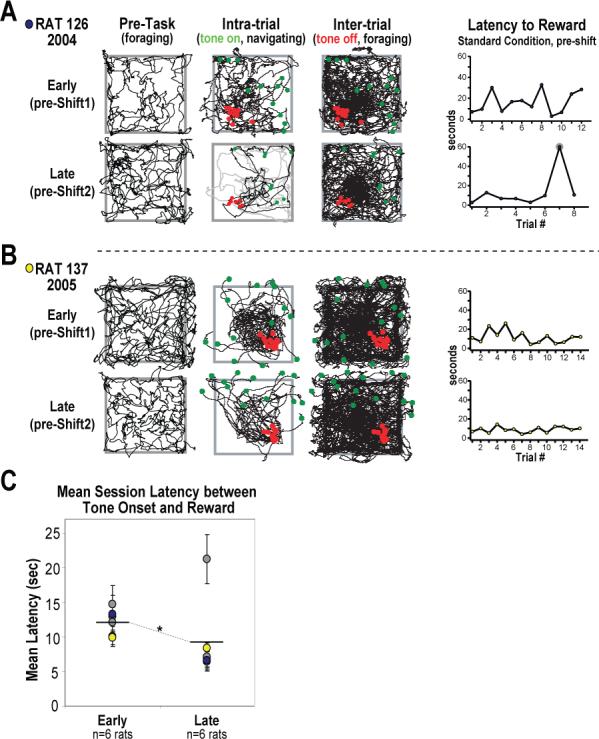
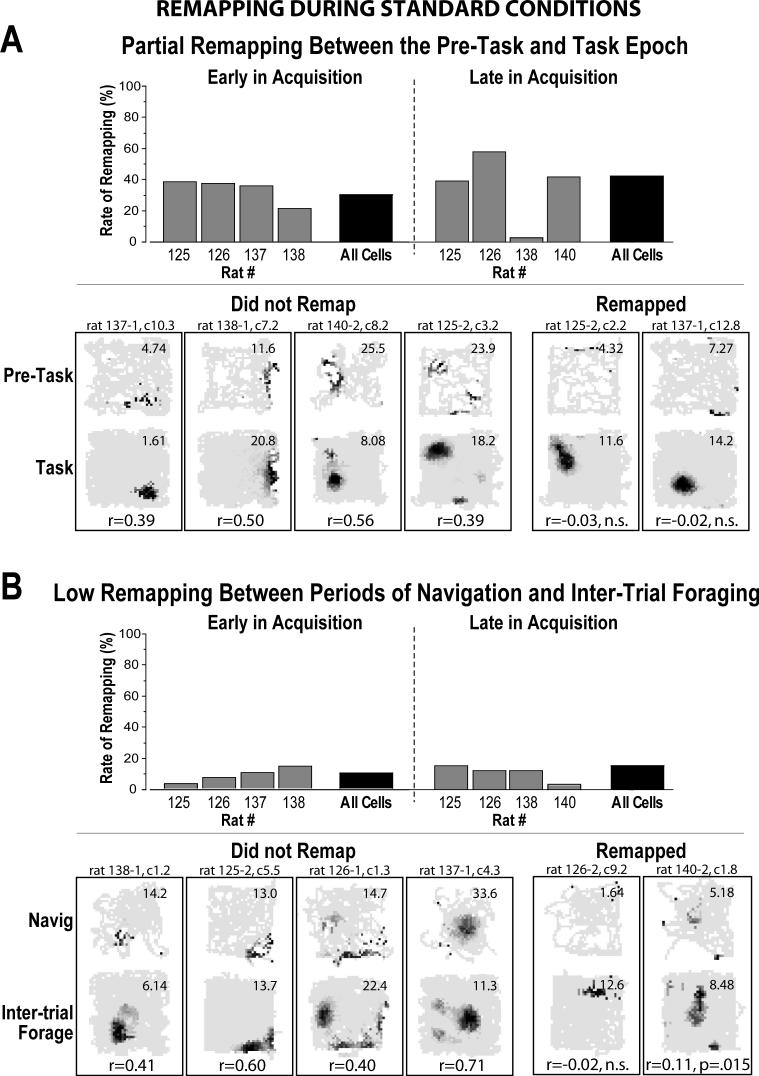
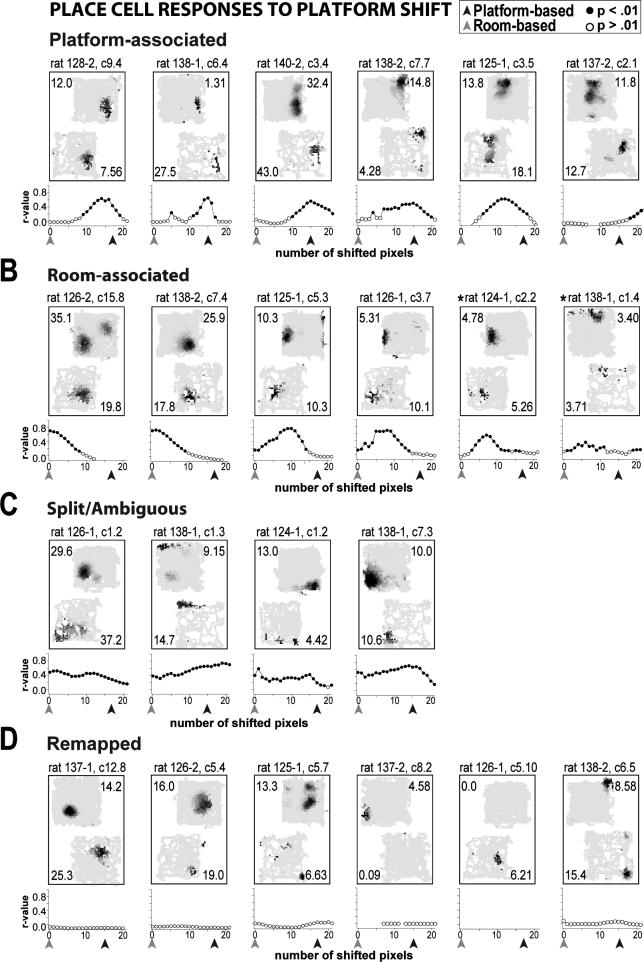
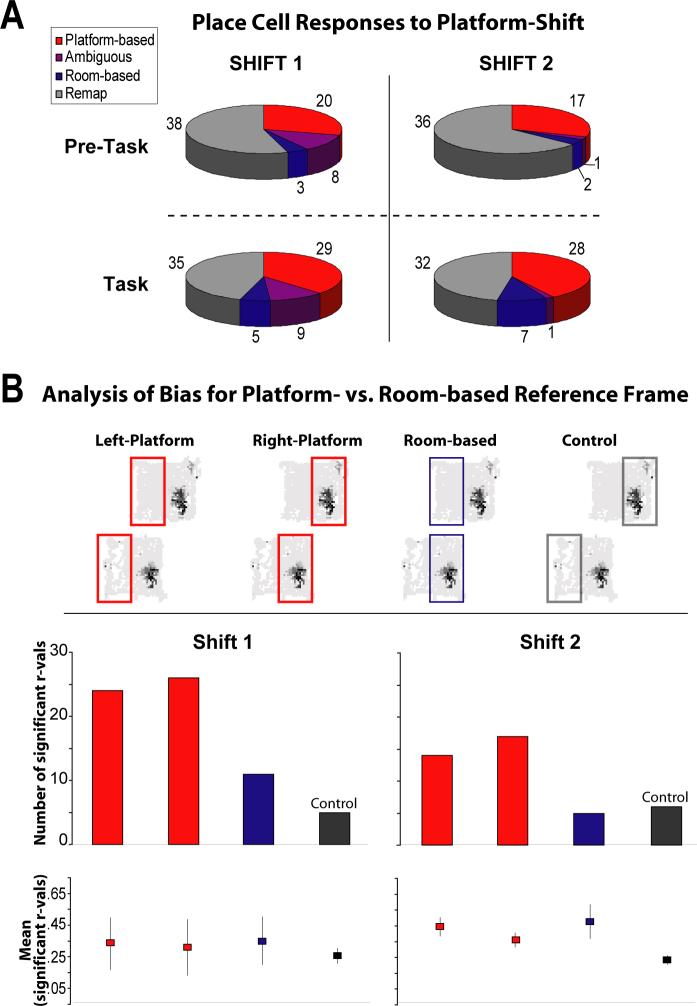
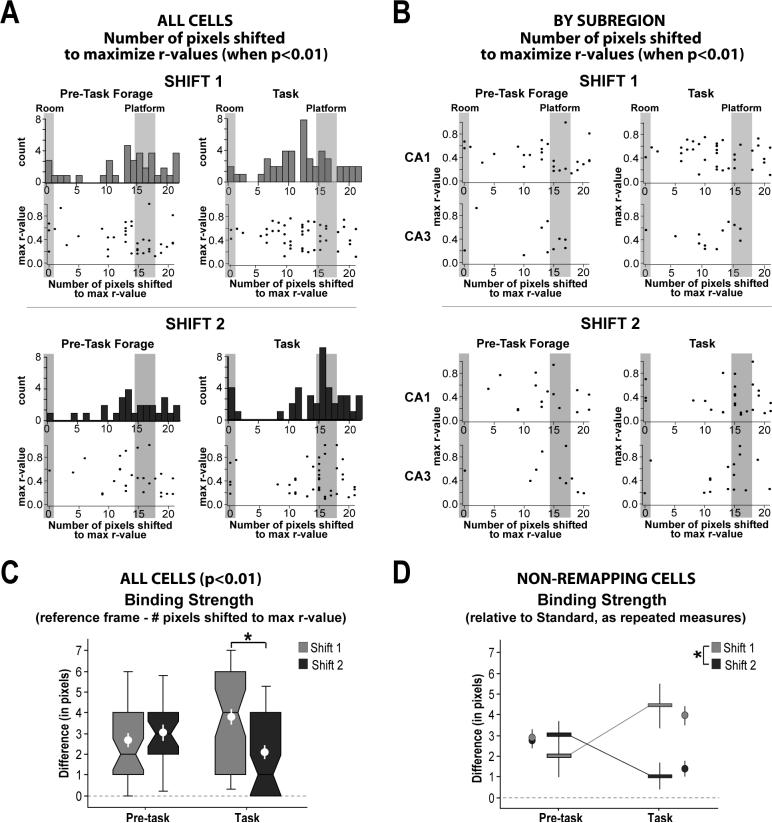
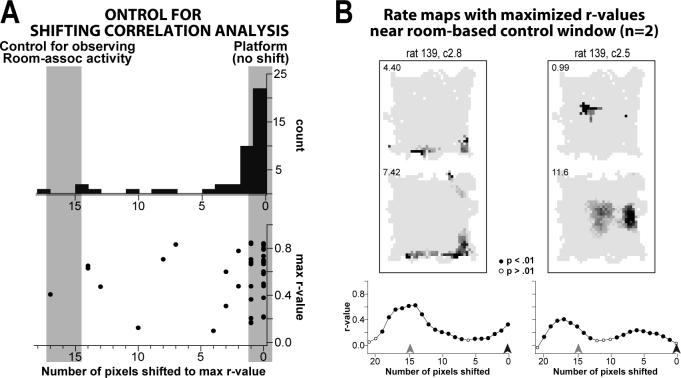
The place-specific activity of hippocampal cells provides downstream structures with information regarding an animal's position within an environment and, perhaps, the location of goals within that environment. In rodents, recent research has suggested that distal cues primarily set the orientation of the spatial representation, whereas the boundaries of the behavioral apparatus determine the locations of place activity. The current study was designed to address possible biases in some previous research that may have minimized the likelihood of observing place activity bound to distal cues. Hippocampal single-unit activity was recorded from six freely moving rats as they were trained to perform a tone-initiated place-preference task on an open-field platform. To investigate whether place activity was bound to the room- or platform-based coordinate frame (or both), the platform was translated within the room at an "early" and at a "late" phase of task acquisition (Shift 1 and Shift 2). At both time points, CA1 and CA3 place cells demonstrated room-associated and/or platform-associated activity, or remapped in response to the platform shift. Shift 1 revealed place activity that reflected an interaction between a dominant platform-based (proximal) coordinate frame and a weaker room-based (distal) frame because many CA1 and CA3 place fields shifted to a location intermediate to the two reference frames. Shift 2 resulted in place activity that became more strongly bound to either the platform- or room-based coordinate frame, suggesting the emergence of two independent spatial frames of reference (with many more cells participating in platform-based than in room-based representations).
Figures
References
-
- Anderson MI, Killing S, Morris C, O'Donoghue A, Onyiagha D, Stevenson R, Verriotis M, Jeffery KJ. Behavioral correlates of the distributed coding of spatial context. Hippocampus. 2006;16:730–742. - PubMed
-
- Blodgett HC, McCutchan K, Matthews R. Spatial learning in the T-maze; the influence of direction, turn, and food location. J Exp Psychol. 1949;39:800–809. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous