

Multifunctional role of protein kinase C in regulating the formation and maturation of specific synapses
- PMID: 17959813
- PMCID: PMC6673217
- DOI: 10.1523/JNEUROSCI.3305-07.2007
Multifunctional role of protein kinase C in regulating the formation and maturation of specific synapses
Abstract
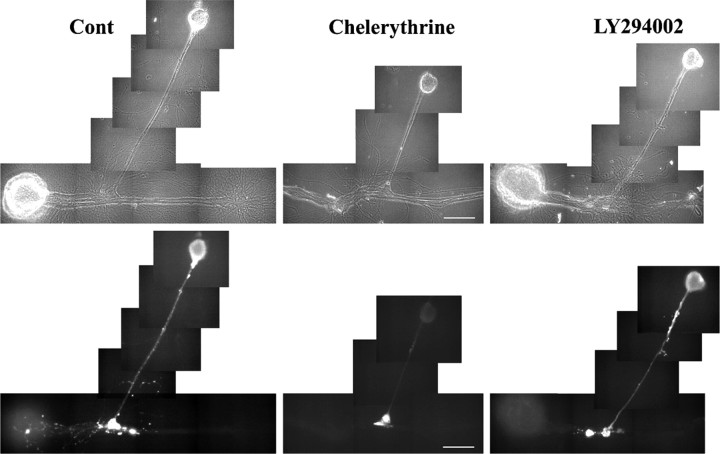
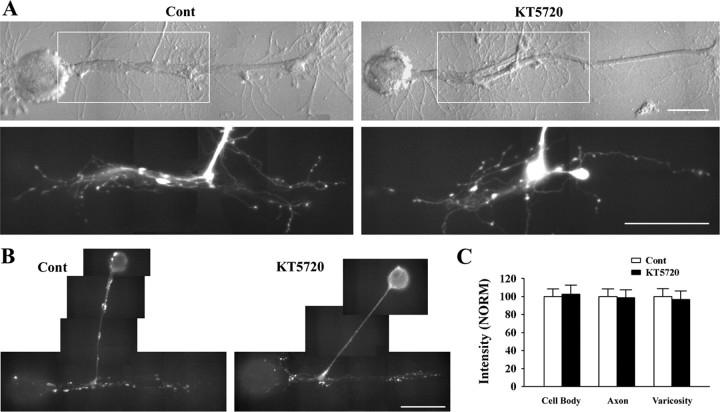
Target-dependent increases in axon growth and varicosities accompany the formation of functional synapses between Aplysia sensory neurons and specific postsynaptic neurons (L7 and not L11). The enhanced growth is regulated in part by a target-dependent increase in the secretion of sensorin, the sensory neuron neuropeptide. We report here that protein kinase C (PKC) activity is required for synapse formation by sensory neurons with L7 and for the target-dependent increases in sensorin synthesis and secretion. Blocking PKC activity reversibly blocked synapse formation and axon growth of sensory neurons contacting L7, but did not affect axon growth of sensory neurons contacting L11 or axon growth of the postsynaptic targets. Blocking PKC activity also blocked the target-induced increase in sensorin synthesis and secretion. Sensorin then activates additional signaling pathways required for synapse maturation and synapse-associated growth. Exogenous anti-sensorin antibody blocked target-induced activation and translocation into sensory neuron nuclei of p42/44 mitogen-activated protein kinase (MAPK), attenuated synapse maturation, and curtailed growth of sensory neurons contacting L7, but not the growth of sensory neurons contacting L11. Inhibitors of MAPK or phosphoinositide 3-kinase also attenuated synapse maturation and curtailed growth and varicosity formation of sensory neurons contacting L7, but not growth of sensory neurons contacting L11. These results suggest that PKC activity regulated by specific cell-cell interactions initiates the formation of specific synapses and the subsequent synthesis and release of a neuropeptide to activate additional signaling pathways required for synapse maturation.
Figures









Similar articles
-
Aplysia cell adhesion molecule and a novel protein kinase C activity in the postsynaptic neuron are required for presynaptic growth and initial formation of specific synapses.J Neurosci. 2010 Jun 23;30(25):8353-66. doi: 10.1523/JNEUROSCI.0546-10.2010. J Neurosci. 2010. PMID: 20573882 Free PMC article.
-
Target-dependent release of a presynaptic neuropeptide regulates the formation and maturation of specific synapses in Aplysia.J Neurosci. 2004 Nov 3;24(44):9933-43. doi: 10.1523/JNEUROSCI.3329-04.2004. J Neurosci. 2004. PMID: 15525778 Free PMC article.
-
Protein kinase C regulates local synthesis and secretion of a neuropeptide required for activity-dependent long-term synaptic plasticity.J Neurosci. 2007 Aug 15;27(33):8927-39. doi: 10.1523/JNEUROSCI.2322-07.2007. J Neurosci. 2007. PMID: 17699674 Free PMC article.
-
Serotonin regulates the secretion and autocrine action of a neuropeptide to activate MAPK required for long-term facilitation in Aplysia.Neuron. 2004 Aug 5;43(3):373-85. doi: 10.1016/j.neuron.2004.07.011. Neuron. 2004. PMID: 15294145
-
Postsynaptic regulation of the development and long-term plasticity of Aplysia sensorimotor synapses in cell culture.J Neurobiol. 1994 Jun;25(6):666-93. doi: 10.1002/neu.480250608. J Neurobiol. 1994. PMID: 8071666 Review.
Cited by
-
Molecular and cellular mechanism of okadaic acid (OKA)-induced neurotoxicity: a novel tool for Alzheimer's disease therapeutic application.Mol Neurobiol. 2014 Dec;50(3):852-65. doi: 10.1007/s12035-014-8699-4. Epub 2014 Apr 8. Mol Neurobiol. 2014. PMID: 24710687 Review.
-
Synapse- and stimulus-specific local translation during long-term neuronal plasticity.Science. 2009 Jun 19;324(5934):1536-40. doi: 10.1126/science.1173205. Epub 2009 May 14. Science. 2009. PMID: 19443737 Free PMC article.
-
Effects of axotomy on cultured sensory neurons of Aplysia: long-term injury-induced changes in excitability and morphology are mediated by different signaling pathways.J Neurophysiol. 2008 Dec;100(6):3209-24. doi: 10.1152/jn.90539.2008. Epub 2008 Oct 8. J Neurophysiol. 2008. PMID: 18842953 Free PMC article.
-
Neuroprotective Effects of Guanosine in Ischemic Stroke-Small Steps towards Effective Therapy.Int J Mol Sci. 2021 Jun 27;22(13):6898. doi: 10.3390/ijms22136898. Int J Mol Sci. 2021. PMID: 34199004 Free PMC article. Review.
-
Aplysia cell adhesion molecule and a novel protein kinase C activity in the postsynaptic neuron are required for presynaptic growth and initial formation of specific synapses.J Neurosci. 2010 Jun 23;30(25):8353-66. doi: 10.1523/JNEUROSCI.0546-10.2010. J Neurosci. 2010. PMID: 20573882 Free PMC article.
References
-
- Aguado F, Carmona MA, Pozas E, Aguilo A, Martinez-Guijarro FJ, Alcantara S, Borrell V, Yuste R, Ibanez CF, Soriano E. BDNF regulates spontaneous correlated activity at early developmental stages by increasing synaptogenesis and expression of the K+/Cl- co-transporter KCC2. Development. 2003;130:1267–1280. - PubMed
-
- Akins MR, Biederer T. Cell-cell interactions in synaptogenesis. Curr Opin Neurobiol. 2006;16:83–89. - PubMed
-
- Alsina B, Vu T, Cohen-Cory S. Visualizing synapse formation in arborizing optic axons in vivo: dynamics and modulation by BDNF. Nat Neurosci. 2001;4:1093–1101. - PubMed
-
- Bailey CH, Kaang BK, Chen M, Martin KC, Lim CS, Casadio A, Kandel ER. Mutation in the phosphorylation sites of MAP kinase blocks learning-related internalization of apCAM in Aplysia sensory neurons. Neuron. 1997;18:913–924. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources