

Auditory cortex mapmaking: principles, projections, and plasticity
- PMID: 17964251
- PMCID: PMC2412907
- DOI: 10.1016/j.neuron.2007.10.013
Auditory cortex mapmaking: principles, projections, and plasticity
Abstract
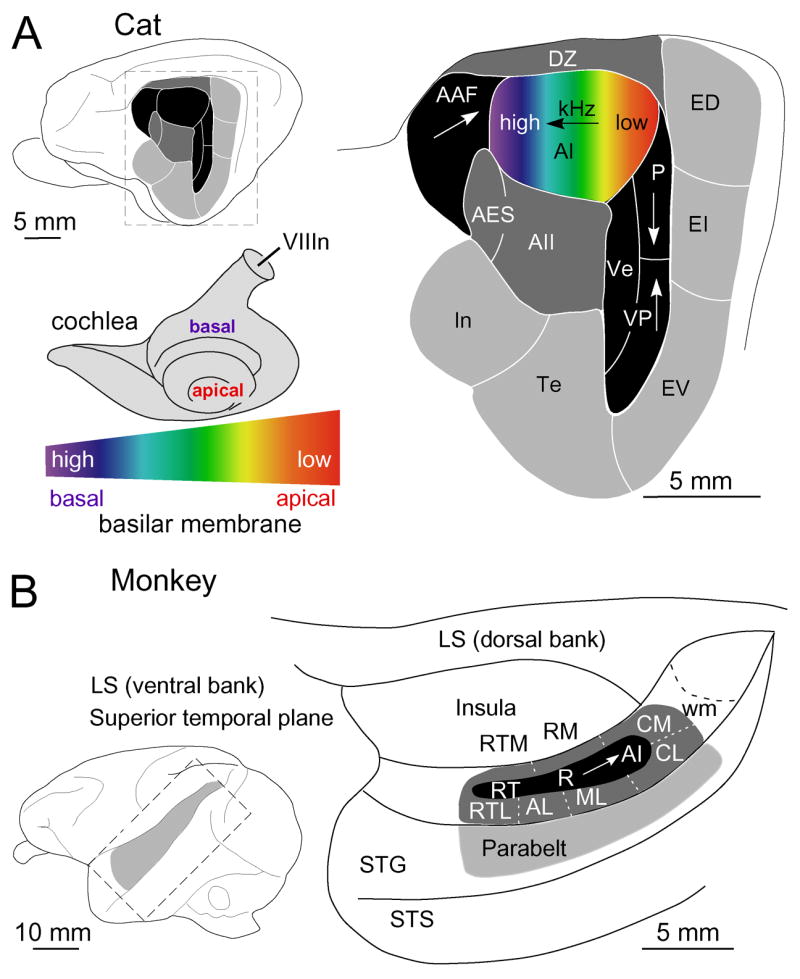
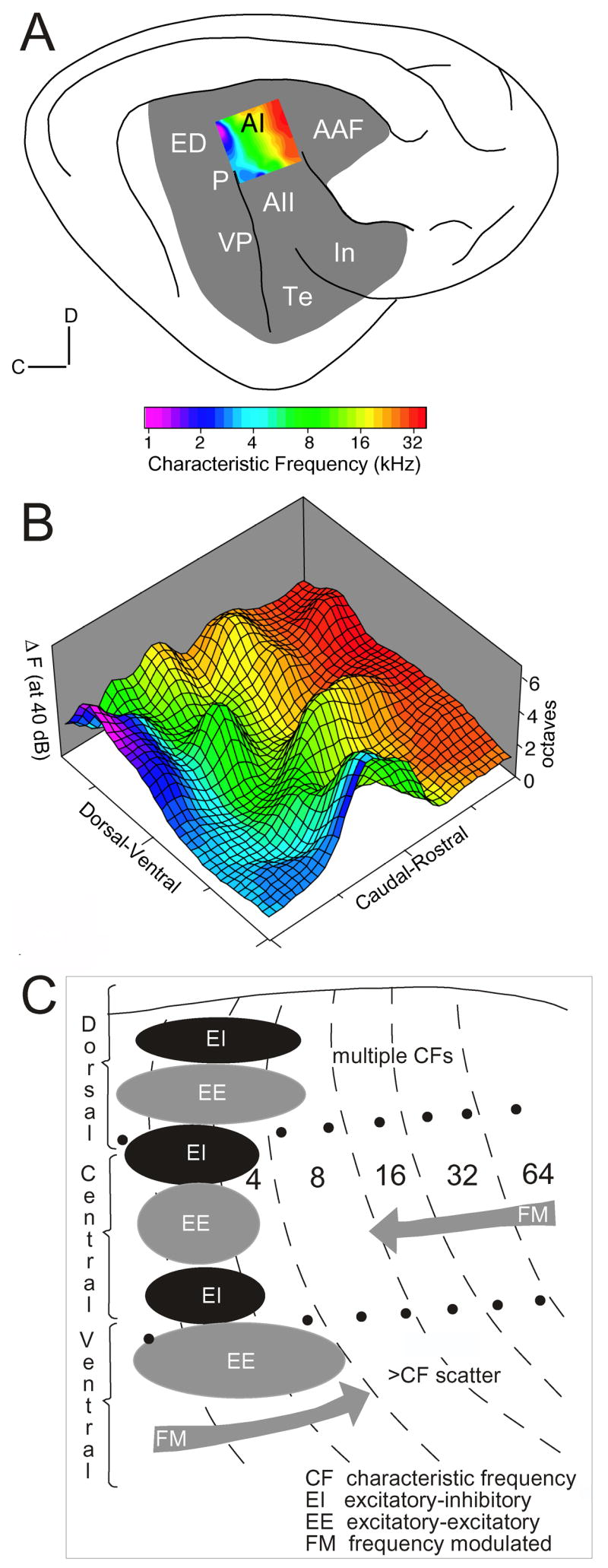
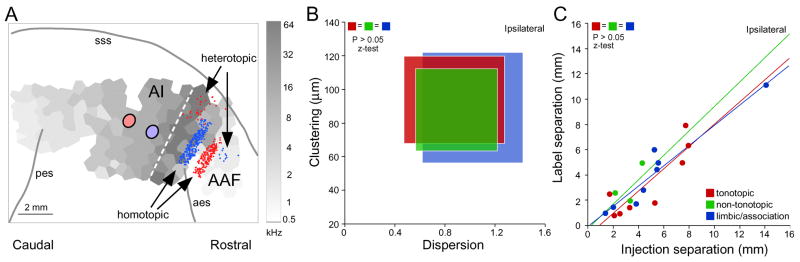
Maps of sensory receptor epithelia and computed features of the sensory environment are common elements of auditory, visual, and somatic sensory representations from the periphery to the cerebral cortex. Maps enhance the understanding of normal neural organization and its modification by pathology and experience. They underlie the derivation of the computational principles that govern sensory processing and the generation of perception. Despite their intuitive explanatory power, the functions of and rules for organizing maps and their plasticity are not well understood. Some puzzles of auditory cortical map organization are that few complete receptor maps are available and that even fewer computational maps are known beyond primary cortical areas. Neuroanatomical evidence suggests equally organized connectional patterns throughout the cortical hierarchy that might underlie map stability. Here, we consider the implications of auditory cortical map organization and its plasticity and evaluate the complementary role of maps in representation and computation from an auditory perspective.
Figures
References
-
- Aleksandrov YI. Learning and memory: traditional and systems approaches. Neurosci Behav Physiol. 2007;36:969–985. - PubMed
-
- Bao S, Chan VT, Merzenich MM. Cortical remodelling induced by activity of ventral tegmental dopamine neurons. Nature. 2001;412:79–83. - PubMed
-
- Bartlett EL, Wang X. Long-lasting modulation by stimulus context in primate auditory cortex. J Neurophysiol. 2005;94:83–104. - PubMed
-
- Bilecen D, Seifritz E, Scheffler K, Scheich H. Amplitopicity of the human auditory cortex: an fMRI study. Neuroimage. 2002;17:701–718. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
