

Functional interdependence at the chromatin level between the MKK6/p38 and IGF1/PI3K/AKT pathways during muscle differentiation
- PMID: 17964260
- PMCID: PMC2693200
- DOI: 10.1016/j.molcel.2007.08.021
Functional interdependence at the chromatin level between the MKK6/p38 and IGF1/PI3K/AKT pathways during muscle differentiation
Abstract
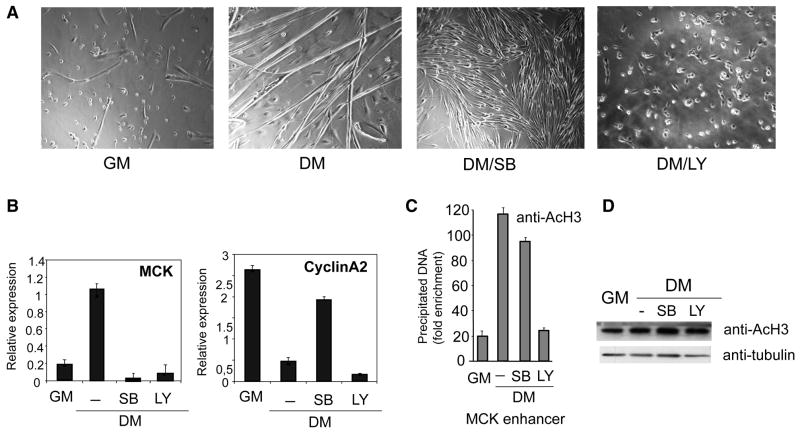
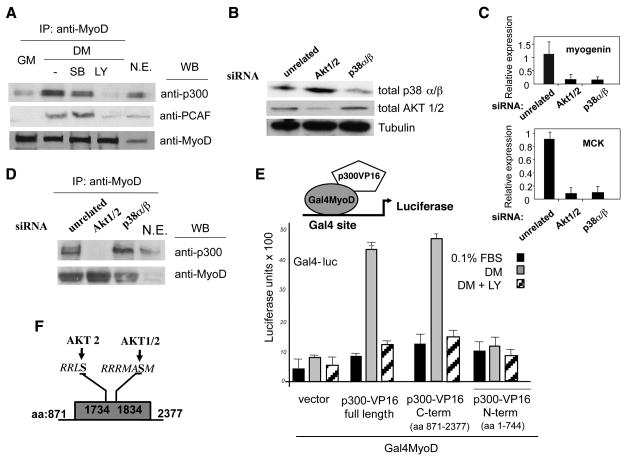
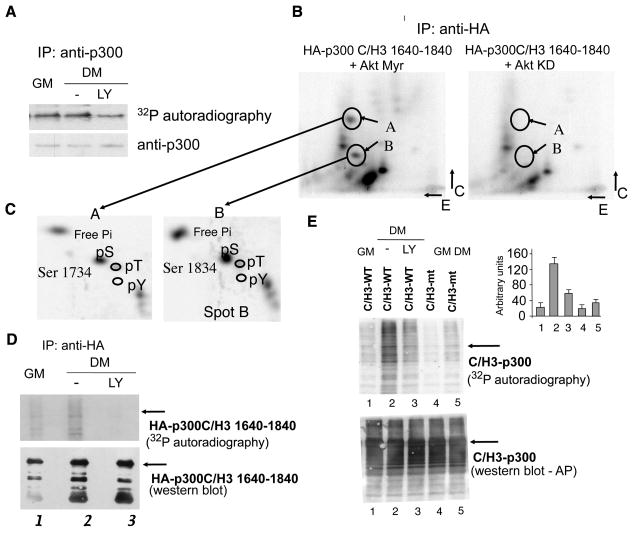
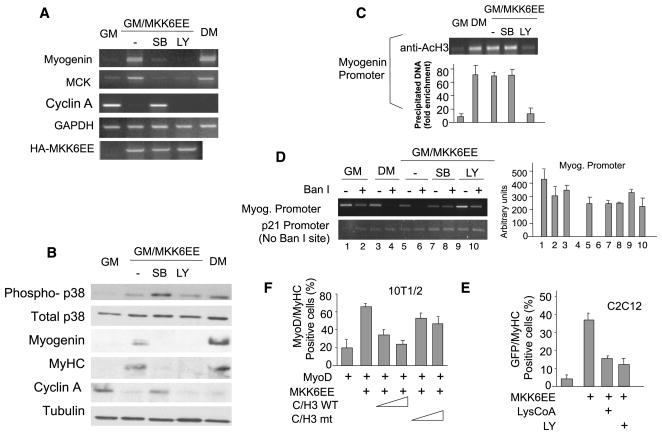
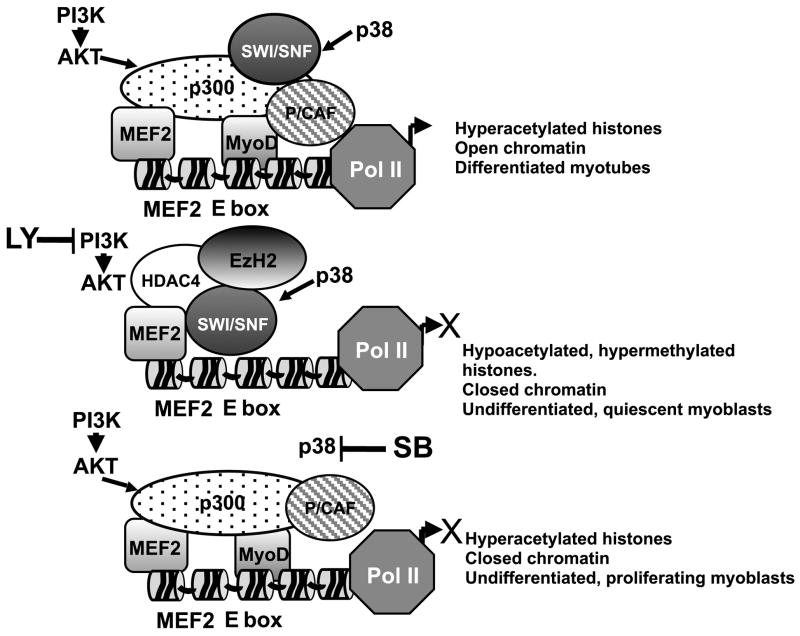
During muscle regeneration, the mechanism integrating environmental cues at the chromatin of muscle progenitors is unknown. We show that inflammation-activated MKK6-p38 and insulin growth factor 1 (IGF1)-induced PI3K/AKT pathways converge on the chromatin of muscle genes to target distinct components of the muscle transcriptosome. p38 alpha/beta kinases recruit the SWI/SNF chromatin-remodeling complex; AKT1 and 2 promote the association of MyoD with p300 and PCAF acetyltransferases, via direct phosphorylation of p300. Pharmacological or genetic interference with either pathway led to partial assembly of discrete chromatin-bound complexes, which reflected two reversible and distinct cellular phenotypes. Remarkably, PI3K/AKT blockade was permissive for chromatin recruitment of MEF2-SWI/SNF complex, whose remodeling activity was compromised in the absence of MyoD and acetyltransferases. The functional interdependence between p38 and IGF1/PI3K/AKT pathways was further established by the evidence that blockade of AKT chromatin targets was sufficient to prevent the activation of the myogenic program triggered by deliberate activation of p38 signaling.
Figures


Similar articles
-
Creatine enhances differentiation of myogenic C2C12 cells by activating both p38 and Akt/PKB pathways.Am J Physiol Cell Physiol. 2007 Oct;293(4):C1263-71. doi: 10.1152/ajpcell.00162.2007. Epub 2007 Jul 25. Am J Physiol Cell Physiol. 2007. PMID: 17652429
-
Myogenic differentiation requires signalling through both phosphatidylinositol 3-kinase and p38 MAP kinase.Cell Signal. 2000 Dec;12(11-12):751-7. doi: 10.1016/s0898-6568(00)00120-0. Cell Signal. 2000. PMID: 11152961
-
TGFβ1-mediated PI3K/Akt and p38 MAP kinase dependent alternative splicing of fibronectin extra domain A in human podocyte culture.Cell Mol Biol (Noisy-le-grand). 2018 Apr 30;64(5):127-135. Cell Mol Biol (Noisy-le-grand). 2018. PMID: 29729706
-
Osteocalcin Induces Proliferation via Positive Activation of the PI3K/Akt, P38 MAPK Pathways and Promotes Differentiation Through Activation of the GPRC6A-ERK1/2 Pathway in C2C12 Myoblast Cells.Cell Physiol Biochem. 2017;43(3):1100-1112. doi: 10.1159/000481752. Epub 2017 Oct 5. Cell Physiol Biochem. 2017. PMID: 28977794
-
Signaling to the chromatin during skeletal myogenesis: novel targets for pharmacological modulation of gene expression.Semin Cell Dev Biol. 2005 Aug-Oct;16(4-5):596-611. doi: 10.1016/j.semcdb.2005.07.005. Semin Cell Dev Biol. 2005. PMID: 16129633 Review.
Cited by
-
Williams Syndrome Transcription Factor is critical for neural crest cell function in Xenopus laevis.Mech Dev. 2012 Sep-Dec;129(9-12):324-38. doi: 10.1016/j.mod.2012.06.001. Epub 2012 Jun 9. Mech Dev. 2012. PMID: 22691402 Free PMC article.
-
Regulation of cellular chromatin state: insights from quiescence and differentiation.Organogenesis. 2010 Jan-Mar;6(1):37-47. doi: 10.4161/org.6.1.11337. Organogenesis. 2010. PMID: 20592864 Free PMC article. Review.
-
Genetic deletion of microRNA biogenesis in muscle cells reveals a hierarchical non-clustered network that controls focal adhesion signaling during muscle regeneration.Mol Metab. 2020 Jun;36:100967. doi: 10.1016/j.molmet.2020.02.010. Epub 2020 Mar 5. Mol Metab. 2020. PMID: 32240622 Free PMC article.
-
Ginsenoside Rb1 and Rb2 upregulate Akt/mTOR signaling-mediated muscular hypertrophy and myoblast differentiation.J Ginseng Res. 2020 May;44(3):435-441. doi: 10.1016/j.jgr.2019.01.007. Epub 2019 Jan 31. J Ginseng Res. 2020. PMID: 32372865 Free PMC article.
-
MicroRNA-106a-5p Inhibited C2C12 Myogenesis via Targeting PIK3R1 and Modulating the PI3K/AKT Signaling.Genes (Basel). 2018 Jul 2;9(7):333. doi: 10.3390/genes9070333. Genes (Basel). 2018. PMID: 30004470 Free PMC article.
References
-
- Berkes CA, Tapscott SJ. MyoD and the transcriptional control of myogenesis. Semin Cell Dev Biol. 2005;16:585–95. - PubMed
-
- Briata P, Forcales SV, Ponassi G, Corte G, Chen C-Y, Karin M, Puri PL, Gherzi R. p38-dependent phosphorylation of the mRNA decay-promoting factor KSRP controls the stability of select myogenic transcripts. Mol Cell. 2005;20(6):891–903. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
