

The essential role of Drosophila HIRA for de novo assembly of paternal chromatin at fertilization
- PMID: 17967064
- PMCID: PMC2041997
- DOI: 10.1371/journal.pgen.0030182
The essential role of Drosophila HIRA for de novo assembly of paternal chromatin at fertilization
Abstract
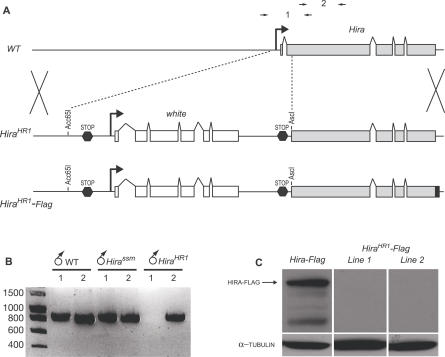
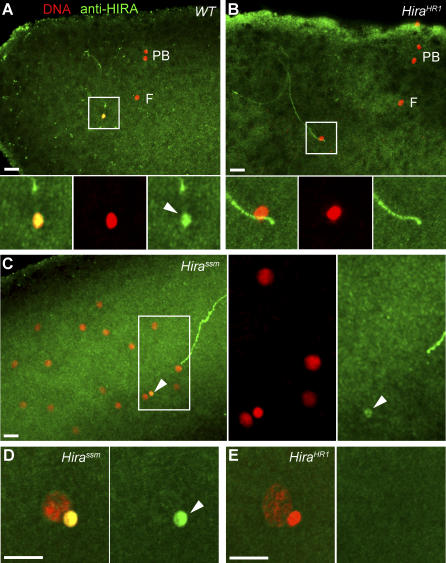
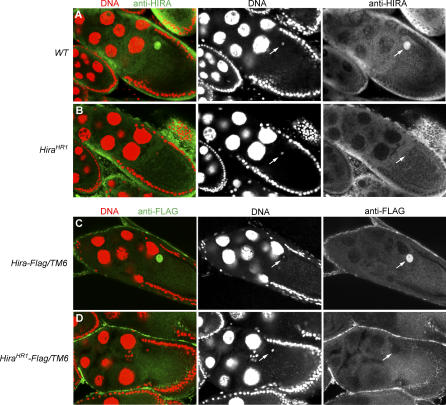
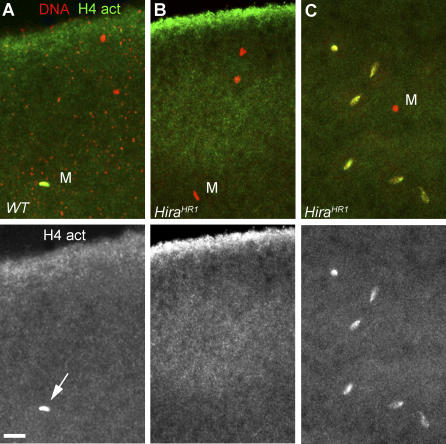
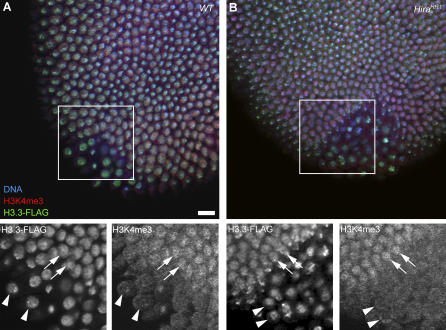
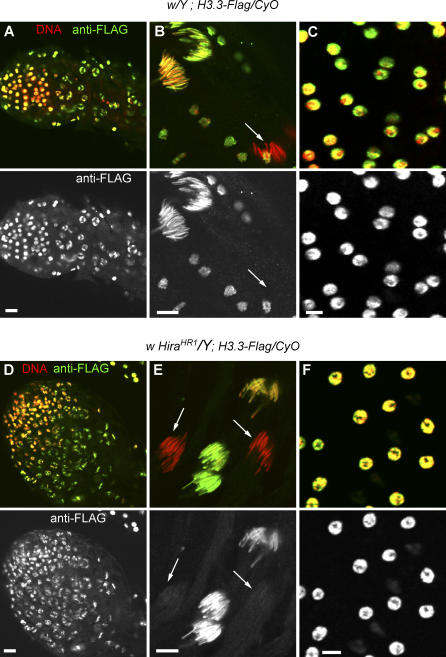
In many animal species, the sperm DNA is packaged with male germ line--specific chromosomal proteins, including protamines. At fertilization, these non-histone proteins are removed from the decondensing sperm nucleus and replaced with maternally provided histones to form the DNA replication competent male pronucleus. By studying a point mutant allele of the Drosophila Hira gene, we previously showed that HIRA, a conserved replication-independent chromatin assembly factor, was essential for the assembly of paternal chromatin at fertilization. HIRA permits the specific assembly of nucleosomes containing the histone H3.3 variant on the decondensing male pronucleus. We report here the analysis of a new mutant allele of Drosophila Hira that was generated by homologous recombination. Surprisingly, phenotypic analysis of this loss of function allele revealed that the only essential function of HIRA is the assembly of paternal chromatin during male pronucleus formation. This HIRA-dependent assembly of H3.3 nucleosomes on paternal DNA does not require the histone chaperone ASF1. Moreover, analysis of this mutant established that protamines are correctly removed at fertilization in the absence of HIRA, thus demonstrating that protamine removal and histone deposition are two functionally distinct processes. Finally, we showed that H3.3 deposition is apparently not affected in Hira mutant embryos and adults, suggesting that different chromatin assembly machineries could deposit this histone variant.
Conflict of interest statement
Competing interests. The authors have declared that no competing interests exist.
Figures
References
-
- Polo SE, Almouzni G. Chromatin assembly: a basic recipe with various flavours. Curr Opin Genet Dev. 2006;16(2):104–111. - PubMed
-
- Akey CW, Luger K. Histone chaperones and nucleosome assembly. Curr Opin Struct Biol. 2003;13(1):6–14. - PubMed
-
- Poccia D, Collas P. Transforming sperm nuclei into male pronuclei in vivo and in vitro. Curr Top Dev Biol. 1996;34:25–88. - PubMed
-
- Wright SJ. Sperm nuclear activation during fertilization. Curr Top Dev Biol. 1999;46:133–178. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
