

Ets-1 is a negative regulator of Th17 differentiation
- PMID: 17967903
- PMCID: PMC2118518
- DOI: 10.1084/jem.20070994
Ets-1 is a negative regulator of Th17 differentiation
Abstract
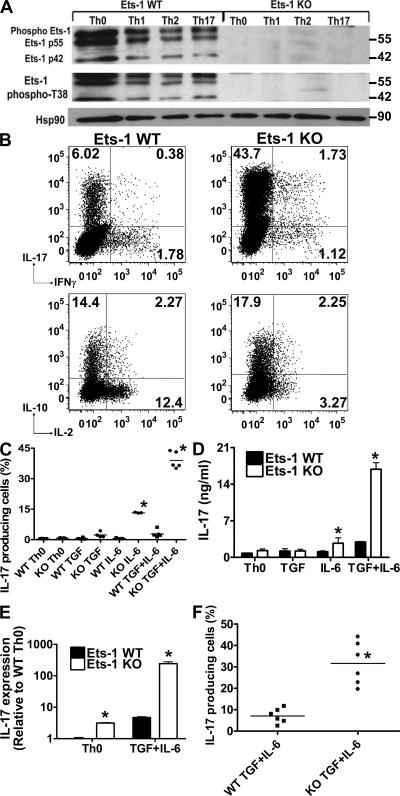
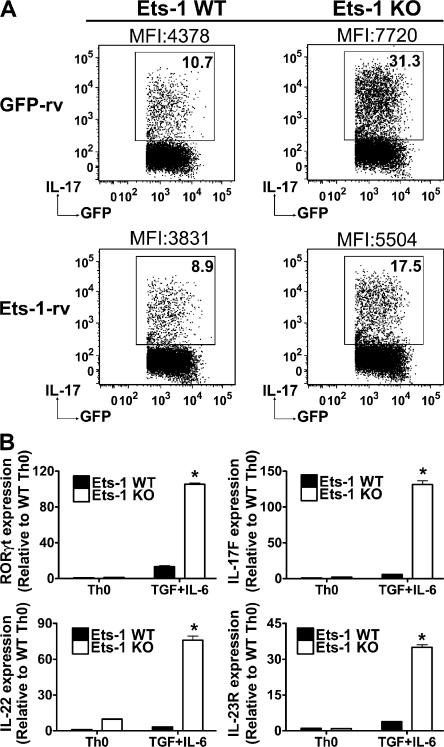
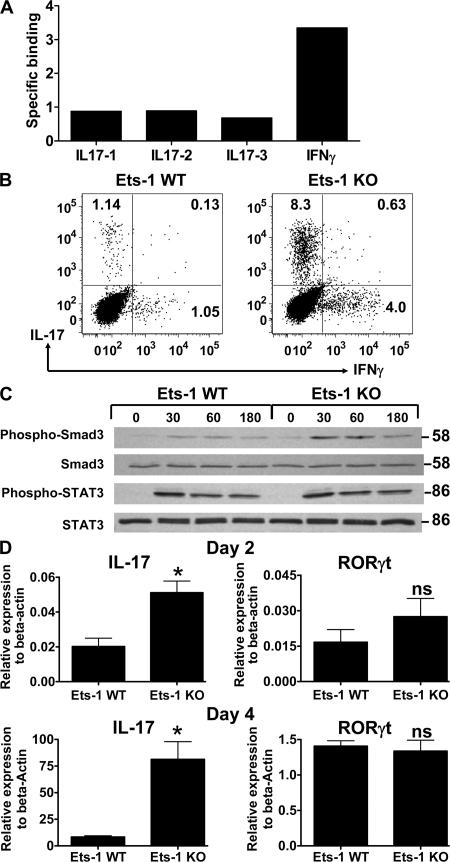
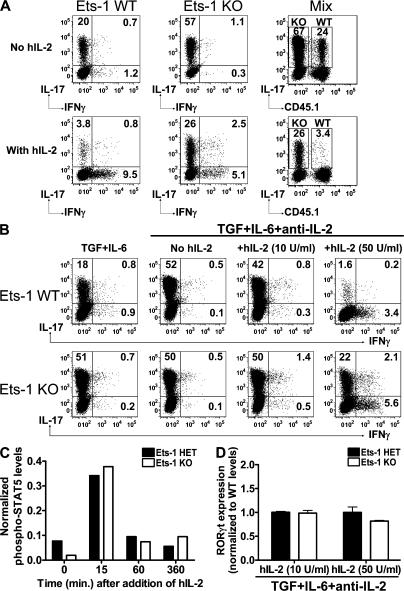
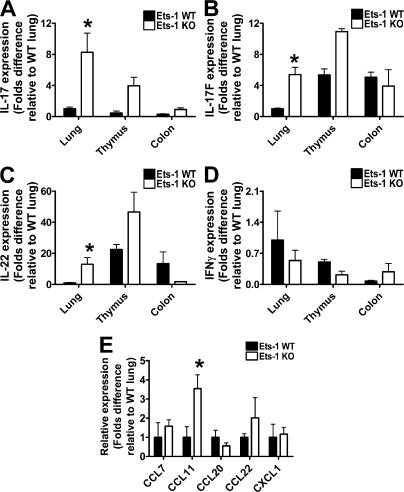
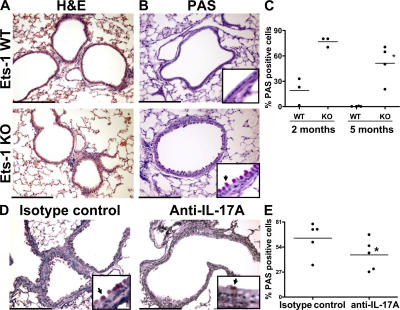
IL-17 is a proinflammatory cytokine that plays a role in the clearance of extracellular bacteria and contributes to the pathology of many autoimmune and allergic conditions. IL-17 is produced mainly by a newly characterized subset of T helper (Th) cells termed Th17. Although the role of Th17 cells in the pathology of autoimmune diseases is well established, the transcription factors regulating the differentiation of Th17 cells remain poorly characterized. We report that Ets-1-deficient Th cells differentiated more efficiently to Th17 cells than wild-type cells. This was attributed to both low IL-2 production and increased resistance to the inhibitory effect of IL-2 on Th17 differentiation. The resistance to IL-2 suppression was caused by a defect downstream of STAT5 phosphorylation, but was not caused by a difference in the level of RORgamma t. Furthermore, Ets-1-deficient mice contained an abnormally high level of IL-17 transcripts in their lungs and exhibited increased mucus production by airway epithelial cells in an IL-17-dependent manner. Based on these observations, we report that Ets-1 is a negative regulator of Th17 differentiation.
Figures
References
-
- Weaver, C.T., R.D. Hatton, P.R. Mangan, and L.E. Harrington. 2007. IL-17 family cytokines and the expanding diversity of effector t cell lineages. Annu. Rev. Immunol. 25:821–852. - PubMed
-
- Ziolkowska, M., A. Koc, G. Luszczykiewicz, K. Ksiezopolska-Pietrzak, E. Klimczak, H. Chwalinska-Sadowska, and W. Maslinski. 2000. High levels of IL-17 in rheumatoid arthritis patients: IL-15 triggers in vitro IL-17 production via cyclosporin A-sensitive mechanism. J. Immunol. 164:2832–2838. - PubMed
-
- Matusevicius, D., P. Kivisakk, B. He, N. Kostulas, V. Ozenci, S. Fredrikson, and H. Link. 1999. Interleukin-17 mRNA expression in blood and CSF mononuclear cells is augmented in multiple sclerosis. Mult. Scler. 5:101–104. - PubMed
-
- Molet, S., Q. Hamid, F. Davoine, E. Nutku, R. Taha, N. Page, R. Olivenstein, J. Elias, and J. Chakir. 2001. IL-17 is increased in asthmatic airways and induces human bronchial fibroblasts to produce cytokines. J. Allergy Clin. Immunol. 108:430–438. - PubMed
-
- Bettelli, E., M. Oukka, and V.K. Kuchroo. 2007. T(H)-17 cells in the circle of immunity and autoimmunity. Nat. Immunol. 8:345–350. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
