

The disulfide relay system of mitochondria is connected to the respiratory chain
- PMID: 17967948
- PMCID: PMC2064786
- DOI: 10.1083/jcb.200707123
The disulfide relay system of mitochondria is connected to the respiratory chain
Abstract
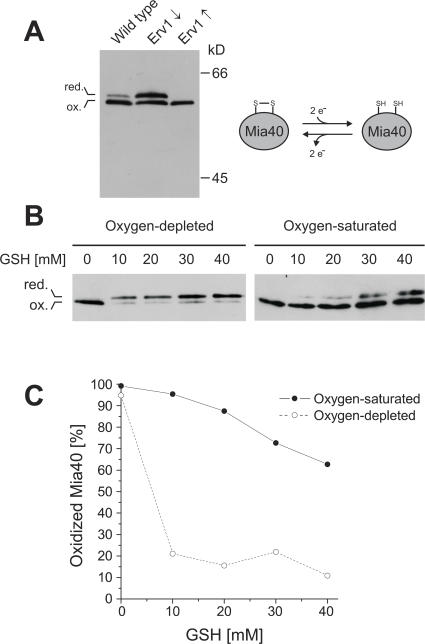
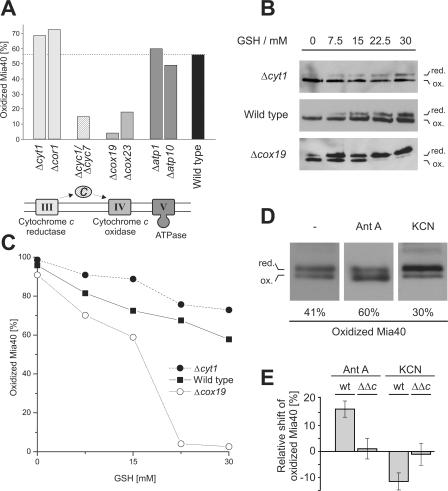
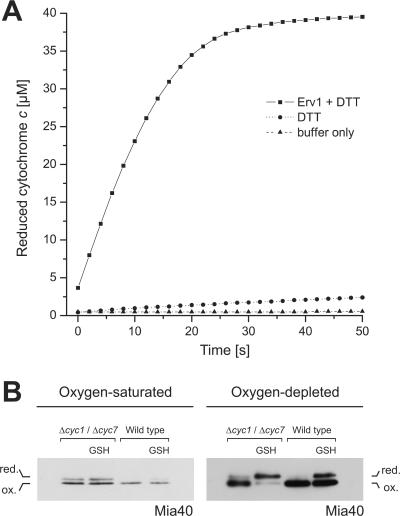
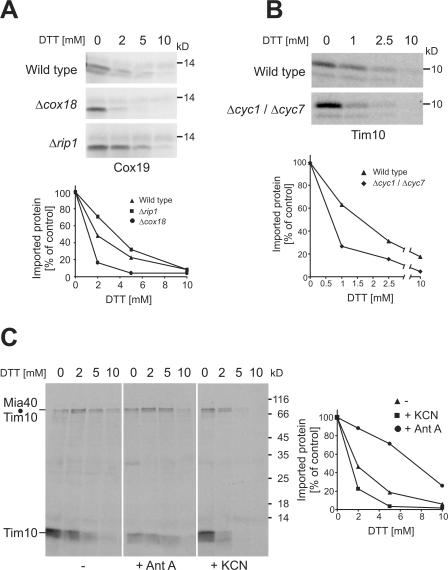
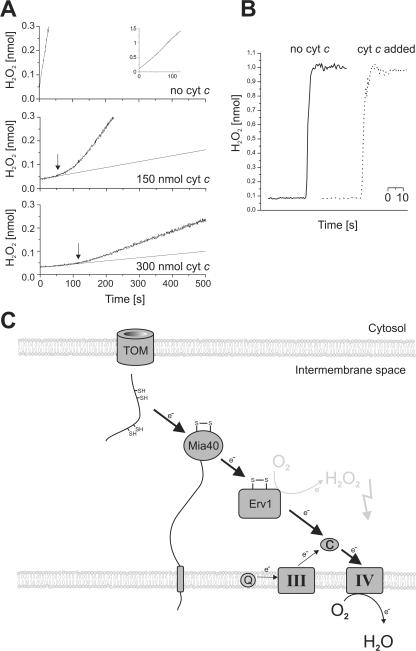
All proteins of the intermembrane space of mitochondria are encoded by nuclear genes and synthesized in the cytosol. Many of these proteins lack presequences but are imported into mitochondria in an oxidation-driven process that relies on the activity of Mia40 and Erv1. Both factors form a disulfide relay system in which Mia40 functions as a receptor that transiently interacts with incoming polypeptides via disulfide bonds. Erv1 is a sulfhydryl oxidase that oxidizes and activates Mia40, but it has remained unclear how Erv1 itself is oxidized. Here, we show that Erv1 passes its electrons on to molecular oxygen via interaction with cytochrome c and cytochrome c oxidase. This connection to the respiratory chain increases the efficient oxidation of the relay system in mitochondria and prevents the formation of toxic hydrogen peroxide. Thus, analogous to the system in the bacterial periplasm, the disulfide relay in the intermembrane space is connected to the electron transport chain of the inner membrane.
Figures
Similar articles
-
Mitochondrial disulfide relay: redox-regulated protein import into the intermembrane space.J Biol Chem. 2012 Feb 10;287(7):4426-33. doi: 10.1074/jbc.R111.270678. Epub 2011 Dec 6. J Biol Chem. 2012. PMID: 22157015 Free PMC article. Review.
-
Structural basis for the disulfide relay system in the mitochondrial intermembrane space.Antioxid Redox Signal. 2010 Nov 1;13(9):1359-73. doi: 10.1089/ars.2010.3099. Antioxid Redox Signal. 2010. PMID: 20136511 Review.
-
The Erv1-Mia40 disulfide relay system in the intermembrane space of mitochondria.Biochim Biophys Acta. 2008 Apr;1783(4):601-9. doi: 10.1016/j.bbamcr.2007.12.005. Epub 2007 Dec 15. Biochim Biophys Acta. 2008. PMID: 18179776 Review.
-
The disulfide relay system of mitochondria is required for the biogenesis of mitochondrial Ccs1 and Sod1.J Mol Biol. 2009 Jan 16;385(2):331-8. doi: 10.1016/j.jmb.2008.10.088. Epub 2008 Nov 7. J Mol Biol. 2009. PMID: 19010334
-
Mitochondrial disulfide bond formation is driven by intersubunit electron transfer in Erv1 and proofread by glutathione.Mol Cell. 2010 Feb 26;37(4):516-28. doi: 10.1016/j.molcel.2010.01.017. Mol Cell. 2010. PMID: 20188670
Cited by
-
Importing mitochondrial proteins: machineries and mechanisms.Cell. 2009 Aug 21;138(4):628-44. doi: 10.1016/j.cell.2009.08.005. Cell. 2009. PMID: 19703392 Free PMC article. Review.
-
Coordinated regulation of sulfur and phospholipid metabolism reflects the importance of methylation in the growth of yeast.Mol Biol Cell. 2011 Nov;22(21):4192-204. doi: 10.1091/mbc.E11-05-0467. Epub 2011 Sep 7. Mol Biol Cell. 2011. PMID: 21900497 Free PMC article.
-
Chemistry and Enzymology of Disulfide Cross-Linking in Proteins.Chem Rev. 2018 Feb 14;118(3):1169-1198. doi: 10.1021/acs.chemrev.7b00123. Epub 2017 Jul 12. Chem Rev. 2018. PMID: 28699750 Free PMC article. Review.
-
Osm1 facilitates the transfer of electrons from Erv1 to fumarate in the redox-regulated import pathway in the mitochondrial intermembrane space.Mol Biol Cell. 2017 Oct 15;28(21):2773-2785. doi: 10.1091/mbc.E16-10-0712. Epub 2017 Aug 16. Mol Biol Cell. 2017. PMID: 28814504 Free PMC article.
-
Mitochondrial copper metabolism and delivery to cytochrome c oxidase.IUBMB Life. 2008 Jul;60(7):421-9. doi: 10.1002/iub.50. IUBMB Life. 2008. PMID: 18459161 Free PMC article. Review.
References
-
- Allen, S., V. Balabanidou, D.P. Sideris, T. Lisowsky, and K. Tokatlidis. 2005. Erv1 mediates the Mia40-dependent protein import pathway and provides a functional link to the respiratory chain by shuttling electrons to cytochrome c. J. Mol. Biol. 353:937–944. - PubMed
-
- Bader, M., W. Muse, D.P. Ballou, C. Gassner, and J.C. Bardwell. 1999. Oxidative protein folding is driven by the electron transport system. Cell. 98:217–227. - PubMed
-
- Bukau, B., J. Weissman, and A. Horwich. 2006. Molecular chaperones and protein quality control. Cell. 125:443–451. - PubMed
-
- Collet, J.F., and J.C. Bardwell. 2002. Oxidative protein folding in bacteria. Mol. Microbiol. 44:1–8. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
