

Drosophila TRP channels require a protein with a distinctive motif encoded by the inaF locus
- PMID: 17968007
- PMCID: PMC2077074
- DOI: 10.1073/pnas.0708368104
Drosophila TRP channels require a protein with a distinctive motif encoded by the inaF locus
Abstract
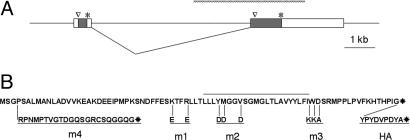
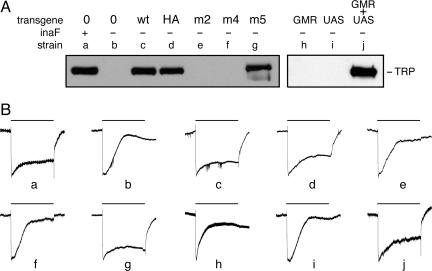
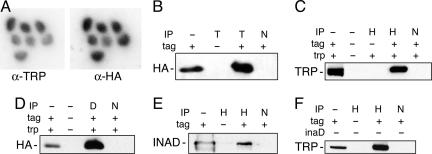
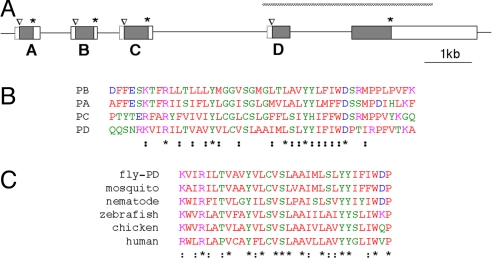
In both vertebrates and invertebrates, ion channels of the TRP superfamily are known to be influenced by a variety of accessory factors, but the list of interacting proteins is acknowledged to be incomplete. Although previous work showed that Drosophila TRP function is disrupted by mutations in the inaF locus, the mechanism of this effect has remained obscure. Here we show that a previously overlooked small protein, INAF-B, is encoded by the locus and fulfills its critical role in retinal physiology. The 81-aa INAF-B gene product is an integral membrane protein that colocalizes to rhabdomeres along with TRP channels. Immunoprecipitation experiments demonstrate that the two proteins participate in a complex, and blotting experiments show that neither protein survives in the absence of the other. Both proteins are normally part of a large supramolecular assembly, the signalplex, but their interaction persists even in the absence of the scaffold for this structure. The inaF locus encodes three other proteins, each of which has diverged from INAF-B except for a 32-aa block of residues that encompasses a transmembrane domain. This conserved sequence defines an inaF motif, representatives of which are found in proteins from organisms as diverse as nematodes, fish, and humans. Given the role of INAF-B, these proteins are good candidates for interacting partners of other members of the TRP superfamily.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
