

FoxO transcription factors suppress Myc-driven lymphomagenesis via direct activation of Arf
- PMID: 17974917
- PMCID: PMC2045131
- DOI: 10.1101/gad.453107
FoxO transcription factors suppress Myc-driven lymphomagenesis via direct activation of Arf
Abstract
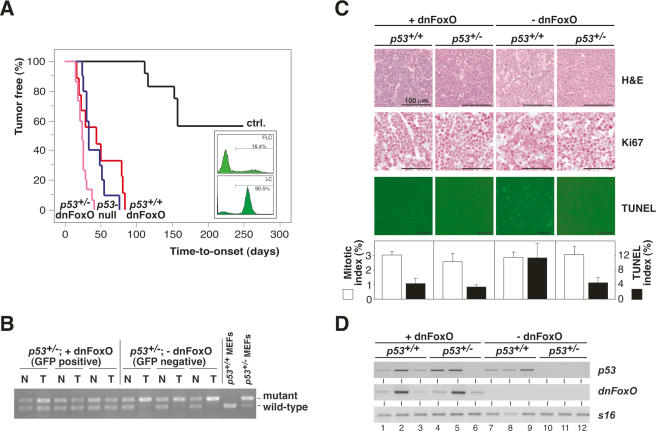
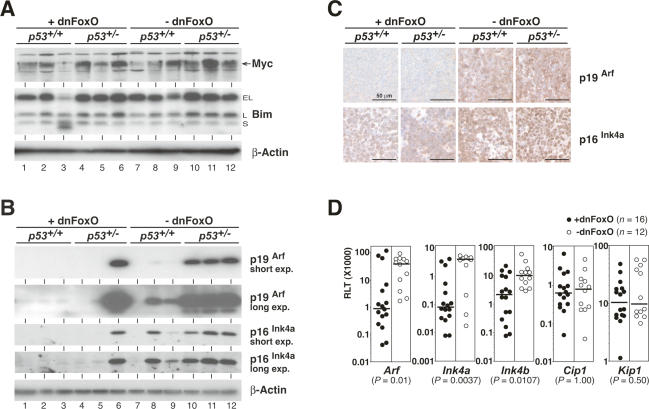
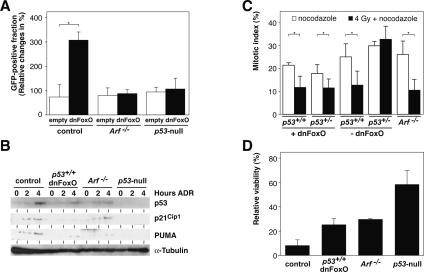
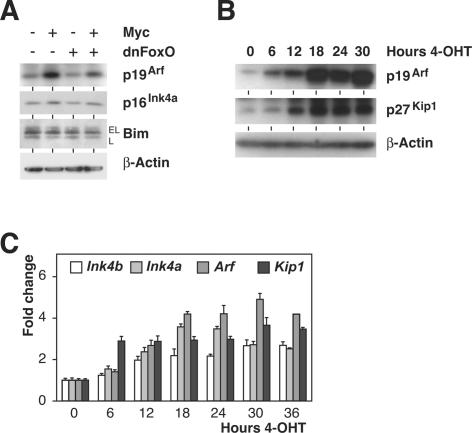
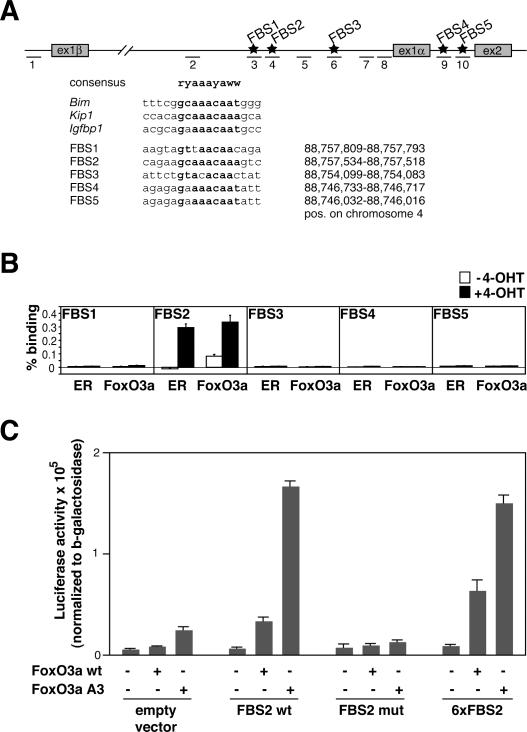
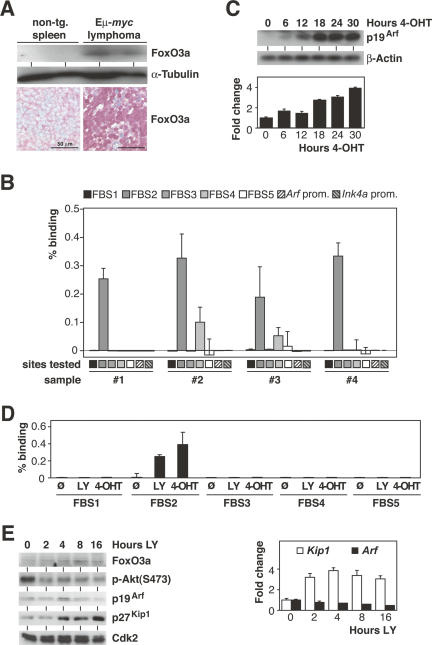
FoxO transcription factors play critical roles in cell cycle control and cellular stress responses, and abrogation of FoxO function promotes focus formation by Myc in vitro. Here we show that stable introduction of a dominant-negative FoxO moiety (dnFoxO) into Emu-myc transgenic hematopoietic stem cells accelerates lymphoma development in recipient mice by attenuating Myc-induced apoptosis. When expressed in Emu-myc; p53(+/-) progenitor cells, dnFoxO alleviates the pressure to inactivate the remaining p53 allele in upcoming lymphomas. Expression of the p53 upstream regulator p19(Arf) is virtually undetectable in most dnFoxO-positive Myc-driven lymphomas. We find that FoxO proteins bind to a distinct site within the Ink4a/Arf locus and activate Arf expression. Moreover, constitutive Myc signaling induces a marked increase in nuclear FoxO levels and stimulates binding of FoxO proteins to the Arf locus. These data demonstrate that FoxO factors mediate Myc-induced Arf expression and provide direct genetic evidence for their tumor-suppressive capacity.
Figures
References
-
- Accili D., Arden K.C., Arden K.C. FoxOs at the crossroads of cellular metabolism, differentiation, and transformation. Cell. 2004;117:421–426. - PubMed
-
- Adams J.M., Harris A.W., Pinkert C.A., Corcoran L.M., Alexander W.S., Cory S., Palmiter R.D., Brinster R.L., Harris A.W., Pinkert C.A., Corcoran L.M., Alexander W.S., Cory S., Palmiter R.D., Brinster R.L., Pinkert C.A., Corcoran L.M., Alexander W.S., Cory S., Palmiter R.D., Brinster R.L., Corcoran L.M., Alexander W.S., Cory S., Palmiter R.D., Brinster R.L., Alexander W.S., Cory S., Palmiter R.D., Brinster R.L., Cory S., Palmiter R.D., Brinster R.L., Palmiter R.D., Brinster R.L., Brinster R.L. The c-myc oncogene driven by immunoglobulin enhancers induces lymphoid malignancy in transgenic mice. Nature. 1985;318:533–538. - PubMed
-
- Bertwistle D., Sherr C.J., Sherr C.J. Regulation of the Arf tumor suppressor in Eμ-Myc transgenic mice: Longitudinal study of Myc-induced lymphomagenesis. Blood. 2006;15:792–794. - PubMed
-
- Birkenkamp K.U., Coffer P.J., Coffer P.J. FOXO transcription factors as regulators of immune homeostasis: Molecules to die for? J. Immunol. 2003;171:1623–1629. - PubMed
-
- Bouchard C., Thieke K., Maier A., Saffrich R., Hanley-Hyde J., Ansorge W., Reed S., Sicinski P., Bartek J., Eilers M., Thieke K., Maier A., Saffrich R., Hanley-Hyde J., Ansorge W., Reed S., Sicinski P., Bartek J., Eilers M., Maier A., Saffrich R., Hanley-Hyde J., Ansorge W., Reed S., Sicinski P., Bartek J., Eilers M., Saffrich R., Hanley-Hyde J., Ansorge W., Reed S., Sicinski P., Bartek J., Eilers M., Hanley-Hyde J., Ansorge W., Reed S., Sicinski P., Bartek J., Eilers M., Ansorge W., Reed S., Sicinski P., Bartek J., Eilers M., Reed S., Sicinski P., Bartek J., Eilers M., Sicinski P., Bartek J., Eilers M., Bartek J., Eilers M., Eilers M. Direct induction of cyclin D2 by Myc contributes to cell cycle progression and sequestration of p27. EMBO J. 1999;18:5321–5333. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous