

Portable flanking sequences modulate CTL epitope processing
- PMID: 17975674
- PMCID: PMC2045603
- DOI: 10.1172/JCI32047
Portable flanking sequences modulate CTL epitope processing
Abstract
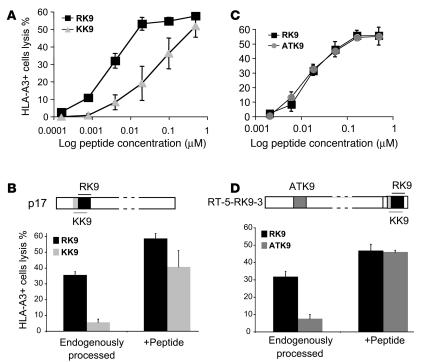
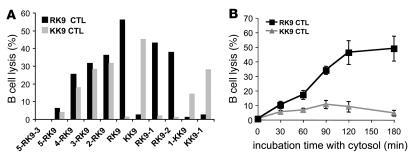
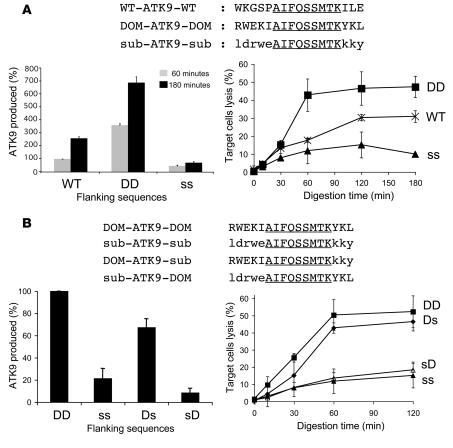
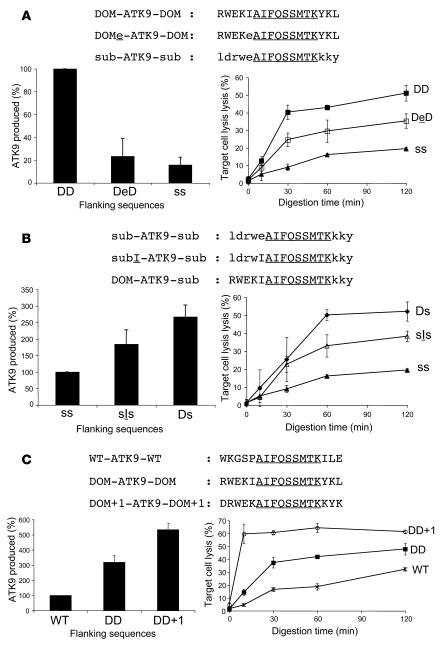
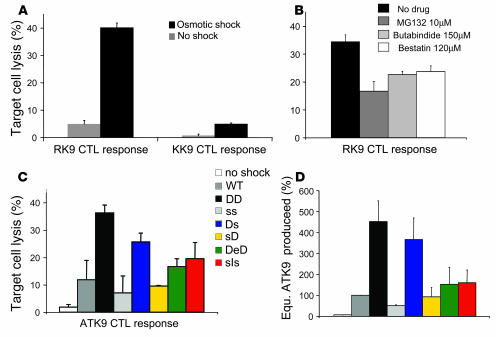
Peptide presentation is critical for immune recognition of pathogen-infected cells by CD8+ T lymphocytes. Although a limited number of immunodominant peptide epitopes are consistently observed in diseases such as HIV-1 infection, the relationship between immunodominance and antigen processing in humans is largely unknown. Here, we have demonstrated that endogenous processing and presentation of a human immunodominant HIV-1 epitope is more efficient than that of a subdominant epitope. Furthermore, we have shown that the regions flanking the immunodominant epitope constitute a portable motif that increases the production and antigenicity of otherwise subdominant epitopes. We used a novel in vitro degradation assay involving cytosolic extracts as well as endogenous intracellular processing assays to examine 2 well-characterized HIV-1 Gag overlapping epitopes presented by the same HLA class I allele, one of which is consistently immunodominant and the other subdominant in infected persons. The kinetics and products of degradation of HIV-1 Gag favored the production of peptides encompassing the immunodominant epitope and destruction of the subdominant one. Notably, cytosolic digestion experiments revealed flanking residues proximal to the immunodominant epitope that increased the production and antigenicity of otherwise subdominant epitopes. Furthermore, specific point mutations in these portable flanking sequences modulated the production and antigenicity of epitopes. Such portable epitope processing determinants provide what we believe is a novel approach to optimizing CTL responses elicited by vaccine vectors.
Figures

Comment in
-
The saga of MHC-bound peptides: a renaissance for antigen presentation?J Clin Invest. 2007 Nov;117(11):3164-6. doi: 10.1172/JCI33997. J Clin Invest. 2007. PMID: 17975658 Free PMC article.
Similar articles
-
Efficient processing of the immunodominant, HLA-A*0201-restricted human immunodeficiency virus type 1 cytotoxic T-lymphocyte epitope despite multiple variations in the epitope flanking sequences.J Virol. 1999 Dec;73(12):10191-8. doi: 10.1128/JVI.73.12.10191-10198.1999. J Virol. 1999. PMID: 10559335 Free PMC article.
-
Selection, transmission, and reversion of an antigen-processing cytotoxic T-lymphocyte escape mutation in human immunodeficiency virus type 1 infection.J Virol. 2004 Jul;78(13):7069-78. doi: 10.1128/JVI.78.13.7069-7078.2004. J Virol. 2004. PMID: 15194783 Free PMC article.
-
CTL epitope distribution patterns in the Gag and Nef proteins of HIV-1 from subtype A infected subjects in Kenya: use of multiple peptide sets increases the detectable breadth of the CTL response.BMC Immunol. 2006 Apr 18;7:8. doi: 10.1186/1471-2172-7-8. BMC Immunol. 2006. PMID: 16620386 Free PMC article.
-
Escape of human immunodeficiency virus from immune control.Annu Rev Immunol. 1997;15:271-96. doi: 10.1146/annurev.immunol.15.1.271. Annu Rev Immunol. 1997. PMID: 9143689 Review.
-
Characteristics of the intrahepatic cytotoxic T lymphocyte response in chronic hepatitis C virus infection.Springer Semin Immunopathol. 1997;19(1):69-83. doi: 10.1007/BF00945026. Springer Semin Immunopathol. 1997. PMID: 9266632 Review.
Cited by
-
Aminopeptidase substrate preference affects HIV epitope presentation and predicts immune escape patterns in HIV-infected individuals.J Immunol. 2012 Jun 15;188(12):5924-34. doi: 10.4049/jimmunol.1200219. Epub 2012 May 14. J Immunol. 2012. PMID: 22586036 Free PMC article.
-
The saga of MHC-bound peptides: a renaissance for antigen presentation?J Clin Invest. 2007 Nov;117(11):3164-6. doi: 10.1172/JCI33997. J Clin Invest. 2007. PMID: 17975658 Free PMC article.
-
Variable fitness impact of HIV-1 escape mutations to cytotoxic T lymphocyte (CTL) response.PLoS Pathog. 2009 Apr;5(4):e1000365. doi: 10.1371/journal.ppat.1000365. Epub 2009 Apr 3. PLoS Pathog. 2009. PMID: 19343217 Free PMC article.
-
HIV-1 gag cytotoxic T lymphocyte epitopes vary in presentation kinetics relative to HLA class I downregulation.J Virol. 2013 Aug;87(15):8726-34. doi: 10.1128/JVI.01040-13. Epub 2013 Jun 5. J Virol. 2013. PMID: 23740989 Free PMC article.
-
HIV-1 p24(gag) derived conserved element DNA vaccine increases the breadth of immune response in mice.PLoS One. 2013;8(3):e60245. doi: 10.1371/journal.pone.0060245. Epub 2013 Mar 28. PLoS One. 2013. PMID: 23555935 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
