

doi: 10.1016/j.jneuroim.2007.09.031.
Epub 2007 Oct 31.
Heterogeneity of EAE mediated by multiple distinct T-effector subsets
Affiliations
- PMID: 17976744
- PMCID: PMC2190732
- DOI: 10.1016/j.jneuroim.2007.09.031
Item in Clipboard
Heterogeneity of EAE mediated by multiple distinct T-effector subsets
J Neuroimmunol.
2007 Dec.
Abstract
Both T(H)1 and T(H)17 lymphocytes are implicated in inducing EAE. In mice lacking IFNgamma, T(H)17 are assumed to be the subset responsible for inflammation induction. Here, we demonstrate that IFNgamma KO mice have two additional effector subsets, one that up-regulates T(H)17-associated pro-inflammatory genes, but does not make IL-17 protein, and a second that utilizes IL-12-related elements of the T(H)1 pathway in an IFNgamma-independent manner. In vivo, these two subsets induce demonstrably different disease. By using homogeneous T cell lines, we can dissect the population of autoimmune effector cells, and demonstrate the multiplicity of pro-inflammatory pathways important in disease processes.
Figures
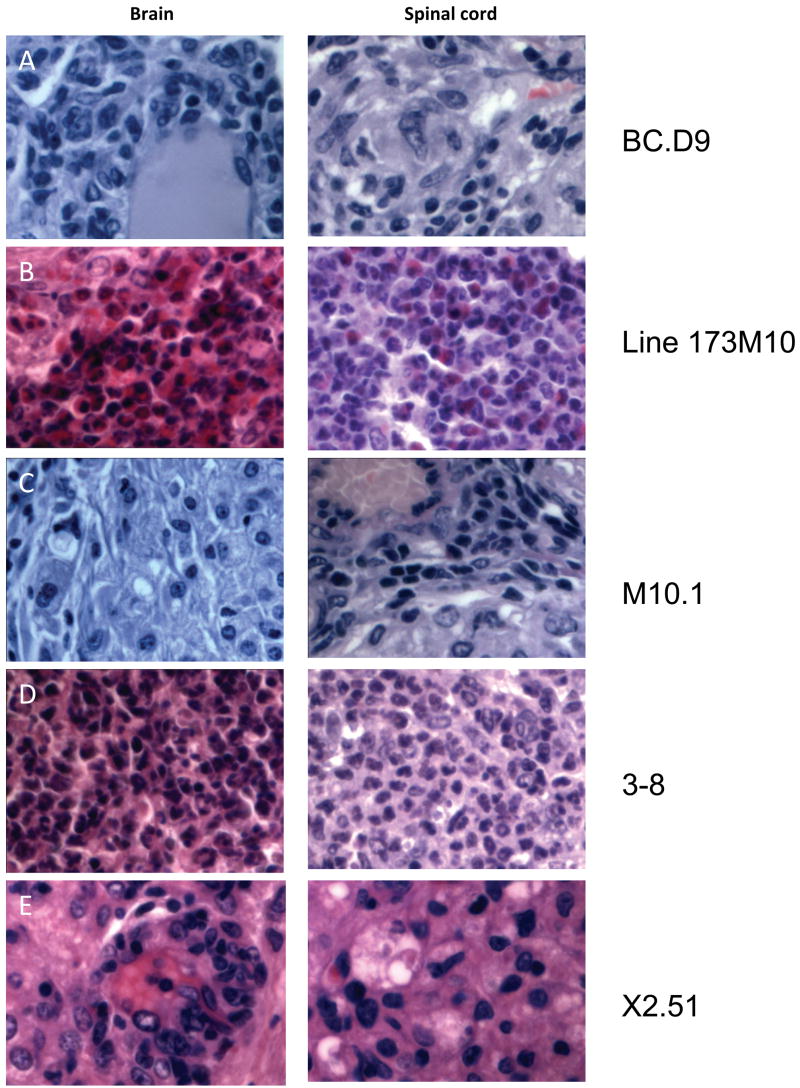
Inflammatory infiltrates in CNS tissues of T cell recipients. Representative hematoxylin and eosin stained sections from brain and spinal cord of indicated T cell clone or line recipients are shown, magnified x600. All mice received 10 × 106 in vitro activated T cells intravenously. Mice were sacrificed within 1–2 days of showing clinical signs of disease. (A) Brain and spinal cord of BCD9 recipient sacrificed 10 days after transfer. Infiltrates consist primarily of macrophages and lymphoid cells, and are concentrated in perivascular and meningeal regions. (B) Brain and spinal cord of line 173M10 recipient 10 days after transfer. Abcesses of neutrophils and eosinophils are present in both brain and spinal cord tissues. (C) Tissues from clone M10.1 recipient 8 days after transfer; macrophages dominate among infiltrating cells. (D) Lateral medulla is completely taken over by infiltrating neutrophils in 3–8 recipients, 10 days after transfer. Spinal cord lesions are rare and isolated, but also consist of very large foci of neutrophils. (E) Tissues from X2.51 recipient 10 days post transfer, dominated by foamy macrophages and lymphoid cells.
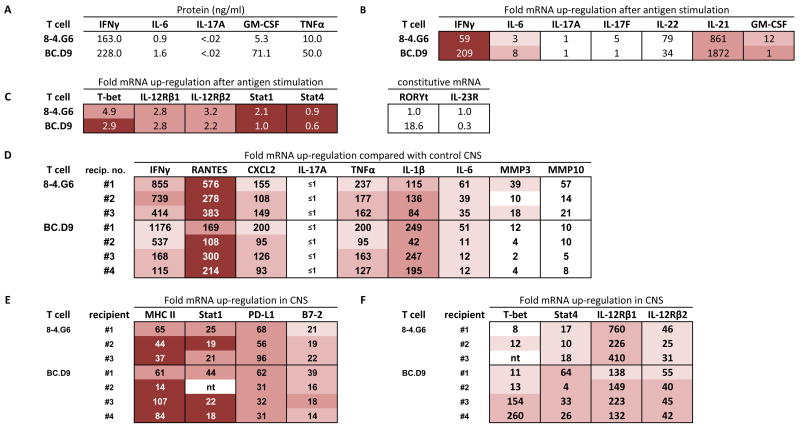
Expression profiles of TH1 cells in vitro (A, B, C) and genes up-regulated in CNS tissue of TH1-induced EAE (D, E, F). (A) ELISA quantitation of supernatants collected 40 hours after activation of T cell clones with MBP peptides (10 μg/ml) and irradiated BALB/c spleen. None of the cytokines were detectable without addition of antigen. (B) Real time PCR analysis of cytokine gene expression 24 hours after activation with antigen and irradiated spleen cells in vitro. mRNA expression is given as fold up-regulation by T cells with antigen compared with T cells incubated in the absence of antigen (both with irradiated antigen presenting cells); a value of 1 indicates that expression after antigen stimulation is the same as expression without antigen. For in vitro cultured cells, this calculated value of fold-up-regulation uses 18S rRNA as housekeeping gene to normalize values. For expression levels, actual Ct values are shown (cDNA from in vitro cultured cells was used at 1/10; Ct of housekeeping gene is nearly identical for each sample shown, within a value of 0.5, and therefore not subtracted). Intensity of shading correlates with intensity of expression level. Lowest Ct values (16–22) are most deeply shaded; three intermediate levels of decreasing shading indicate, respectively, Ct values of 22–26, 26–28, and 28–30. No shading indicates Ct >30. (C) Realtime PCR analysis of up-regulation (after antigen-specific activation, compared with no antigen addition, normalized with 18S rRNA) and expression level (Ct value) of T-bet, IL-12R chains, Stat1, and Stat4. Levels of RORγt and IL-23R did not increase after activation, therefore relative constitutive levels are arbitrarily compared between each T cell and clone 8-4.G6, which had a basal Ct value of 40. (D,E,F) Realtime PCR analysis of gene expression level (Ct value) and fold up-regulation of each gene in CNS of sick mice, compared with expression levels in normal CNS tissue. GAPDH is used as the housekeeping gene for normalization of in vivo tissue. Brain and spinal cord were harvested at first onset of clinical signs of EAE, and cDNA combined in 1:1 ratios for PCR analysis of each BALB/c recipient. Combined cDNA was used at 1/100 dilution.
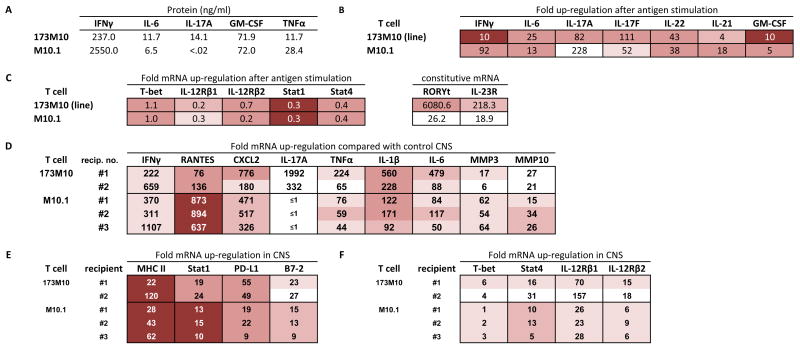
Expression profiles of T cells from a TCR transgenic, IFNγ+ mouse. (A–C) In vitro cultured T cells; see legend for Fig. 2. (D–F) CNS tissue from T cell recipients with EAE; see legend for Fig. 2.
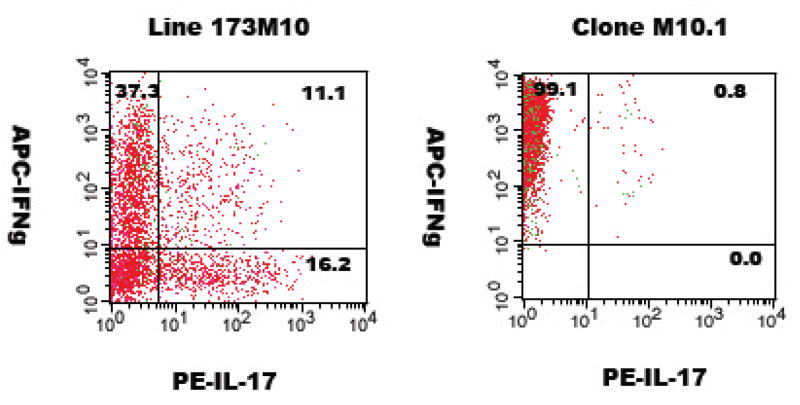
Cytokine analysis by intracellular flow cytometry. T cells were activated by 18 hour incubation with splenic adherent cells and antigen, followed by intracellular cytokine staining for IFNγ and IL-17. Plots shown are gated on CD4+ cells.
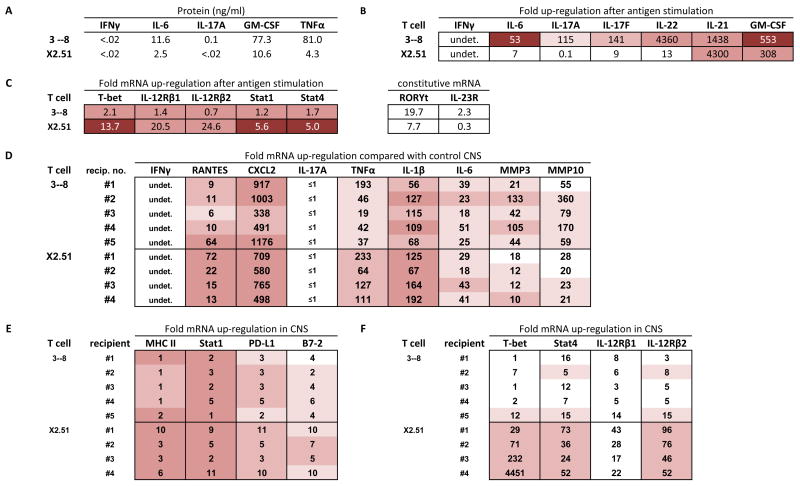
Expression profiles of BALB-GKO-derived T cells. (A-C) In vitro cultured cells; see legend for Fig. 2. (A) Levels of IL-17 protein were undetectable (<.02 ng/ml) in 6 of 9 independently generated supernatants, and present at .08, 0.25, and 0.31 ng/ml in three of nine supernatants (D–F) CNS tissue from sick T cell clone recipients; see legend for Figure 2.
Comment in
-
Many roads lead to Rome: heterogeneity among encephalitogenic T cell clones.J Neuroimmunol. 2007 Dec;192(1-2):1-2. doi: 10.1016/j.jneuroim.2007.10.011. Epub 2007 Nov 26. J Neuroimmunol. 2007. PMID: 18031831 No abstract available.
References
-
- Abromson-Leeman S, Alexander J, Bronson R, Carroll J, Southwood S, Dorf M. Experimental autoimmune encephalomyelitis-resistant mice have highly encephalitogenic myelin basic protein (MBP)-specific T cell clones that recognize a MBP peptide with high affinity for MHC class II. J Immunol. 1995a;154:388–398. - PubMed
-
- Acosta-Rodriguez EV, Rivino L, Geginat J, Jarrossay D, Gattorno M, Lanzavecchia A, Sallusto F, Napolitani G. Surface phenotype and antigenic specificity of human interleukin 17-producing T helper memory cells. Nat Immunol. 2007;8:639–646. - PubMed
-
- Bettelli E, Carrier Y, Gao W, Korn T, Strom TB, Oukka M, Weiner HL, Kuchroo VK. Reciprocal developmental pathways for the generation of pathogenic effector TH17 and regulatory T cells. Nature. 2006;441:235–238. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
