

ATP and cytochrome c-dependent activation of caspase-9 during hypoxia in the cerebral cortex of newborn piglets
- PMID: 17976908
- PMCID: PMC2175014
- DOI: 10.1016/j.neulet.2007.09.072
ATP and cytochrome c-dependent activation of caspase-9 during hypoxia in the cerebral cortex of newborn piglets
Abstract
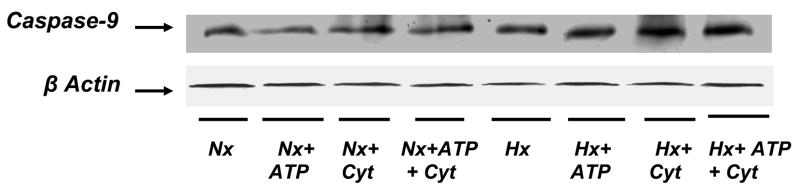
In previous studies, we have shown that cerebral hypoxia results in increased activity of caspase-9, the initiator caspase, and caspase-3, in the cytosolic fraction of the cerebral cortex of newborn piglets. The present study examines the mechanism of caspase-9 activation during hypoxia and tests the hypothesis that the ATP and cytochrome c-dependent activation of caspase-9 increases in the cytosol of the cerebral cortex of newborn piglets. Newborn piglets were divided into normoxic (Nx, n=4), and hypoxic (Hx, n=4) groups. Anesthetized, ventilated animals were exposed to an FiO(2) of 0.21 (Nx) or 0.07 (Hx) for 60 min. Cerebral tissue hypoxia was documented biochemically by determining levels of ATP and phosphocreatine (PCr). Cytosolic fraction was isolated and passed through a G25-Sephadex column to remove endogenous ATP and cytochrome c. Fractions were collected and protein determined by UV spectrophotometry at 280 nm. Eluted high-molecular weight samples from normoxic and hypoxic animals were divided into four subgroups: subgroup 1 (control), incubated without added ATP and cytochrome c; subgroup 2, incubated with added ATP; subgroup 3, incubated with added cytochrome c; and subgroup 4, incubated with added ATP and cytochrome c. The incubation was carried out at 37 degrees C for 30 min. Following incubation, the protein was separated by 12% SDS-PAGE and active caspase-9 was detected using specific active caspase-9 antibody. Protein bands were detected by enhanced chemiluminescence. Protein density was determined by imaging densitometry and expressed as absorbance (OD x mm(2)). ATP (mumol/g brain) level was 4.7 +/- 0.18 in normoxic, as compared to 1.53 +/- 0.16 in hypoxic (p < 0.05 vs. Nx). PCr (mumol/g brain) level was 4.03 +/- 0.11 in the normoxic and 1.1 +/- 0.3 in the hypoxic brain (p < 0.05 vs. Nx). In the normoxic preparations, active caspase-9 density increased by 9, 4 and 20% in the presence of ATP, cytochrome c and ATP+cytochrome c, respectively. In the hypoxic preparations, active caspase-9 density increased by 30, 45 and 60% in the presence of ATP, cytochrome c and ATP+cytochrome c, respectively. These results show that incubation with ATP, cytochrome c and ATP+cytochrome c result in a significantly increased activation of caspase-9 in the hypoxic group (p < 0.05). We conclude that the ATP and cytochrome c dependent activation of caspase-9 is increased during hypoxia. We propose that the ATP and cytochrome c sites of apoptotic protease activating factor I that mediate caspase-9 activation are modified during hypoxia.
Figures

References
-
- Ashraf QM, Haider HS, Katsetos CD, Delivoria-Papadopoulos M, Mishra OP. Nitric oxide mediated alterations of protein tyrosine phosphatase activity and expression during hypoxia in the cerebral cortex of newborn piglets. Neurosci Lett. 2004;362:108–112. - PubMed
-
- Bao Q, Shi Y. Apoptosome: a platform for the activation of initiator caspases. Cell Death Differ. 2007;14:56–65. - PubMed
-
- Chinnaiyan AM, O’Rourke K, Lane BR, Dixit VM. Interaction of CED-4 with CED-3 and CED-9: a molecular framework for cell death. Science. 1997;275:1122–1126. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
