

The peripheral membrane subunits of the SAM complex function codependently in mitochondrial outer membrane biogenesis
- PMID: 17978093
- PMCID: PMC2174179
- DOI: 10.1091/mbc.e07-08-0796
The peripheral membrane subunits of the SAM complex function codependently in mitochondrial outer membrane biogenesis
Abstract
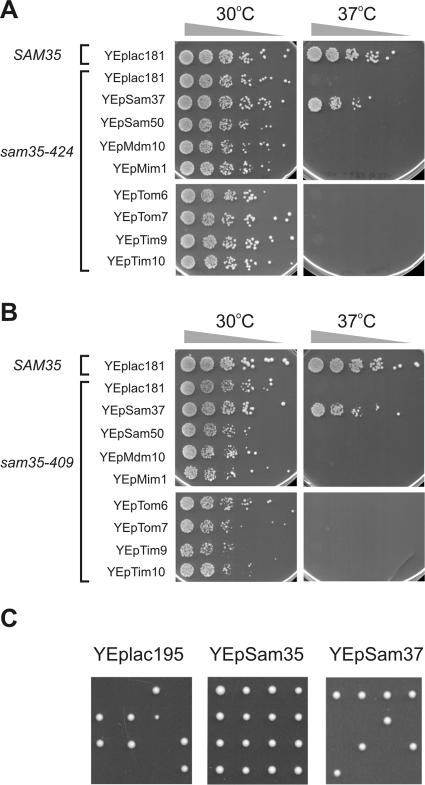
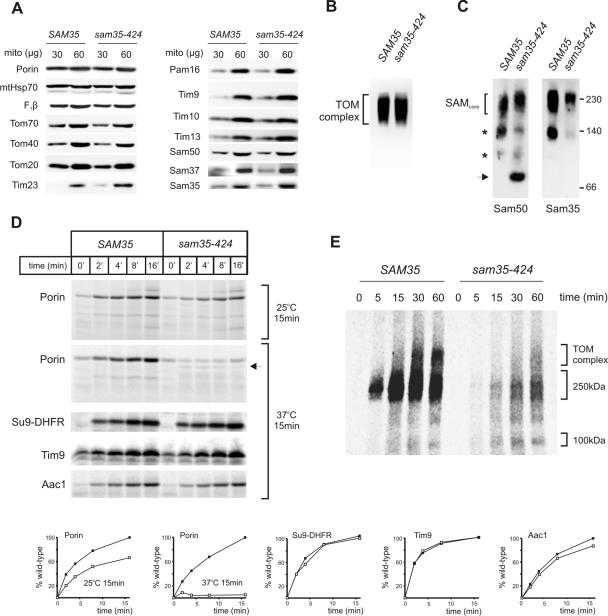
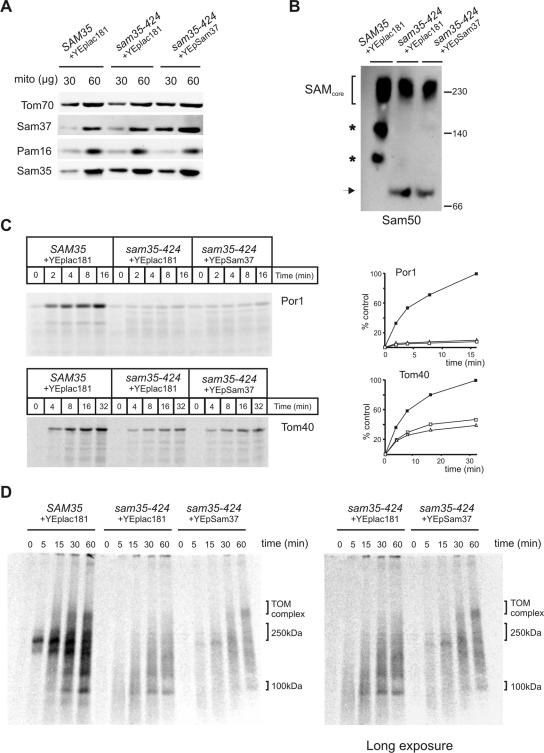
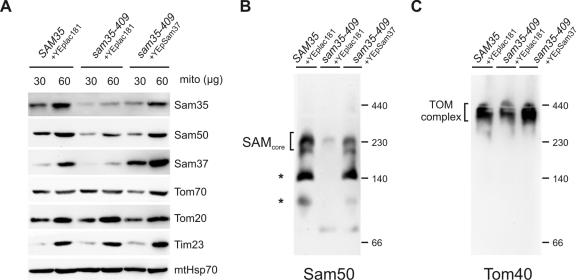
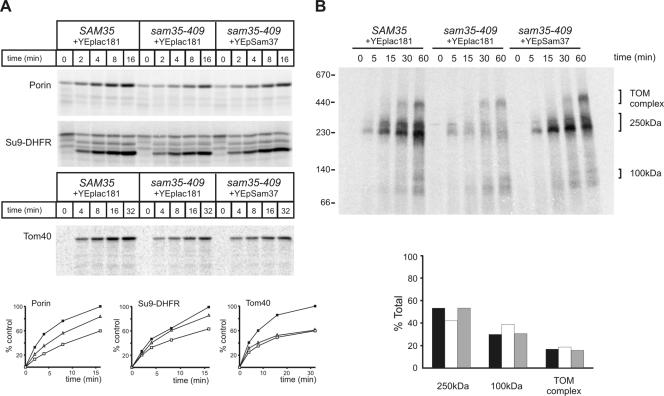
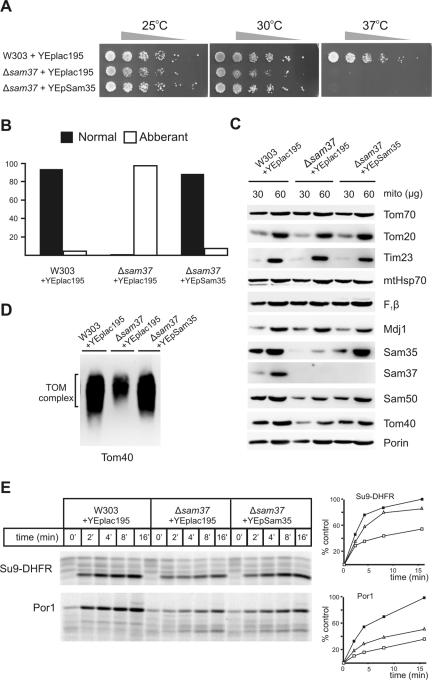
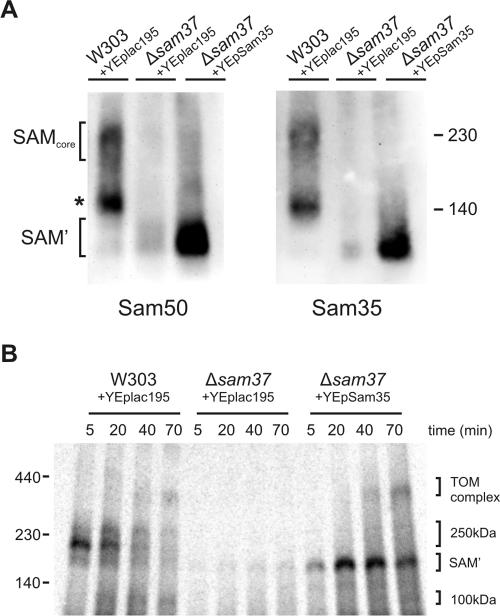
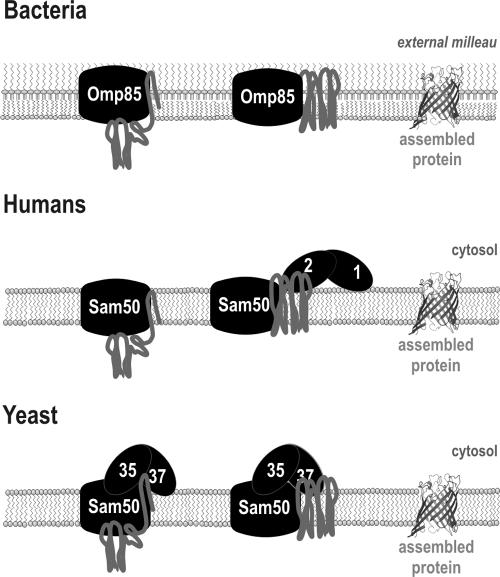
The sorting and assembly machinery (SAM) complex functions in the assembly of beta-barrel proteins into the mitochondrial outer membrane. It is related to the Omp85/YaeT machinery in bacterial outer membranes, but the eukaryotic SAM complex is distinguished by two peripheral subunits, Sam37 and Sam35, that sit on the cytosolic face of the complex. The function of these subunits in beta-barrel protein assembly is currently unclear. By screening a library of sam35 mutants, we show that 13 distinct alleles were each specifically suppressed by overexpression of SAM37. Two of these mutants, sam35-409 and sam35-424, show distinct phenotypes that enable us to distinguish the function of Sam35 from that of Sam37. Sam35 is required for the SAM complex to bind outer membrane substrate proteins: destabilization of Sam35 inhibits substrate binding by Sam50. Sam37 acts later than Sam35, apparently to assist release of substrates from the SAM complex. Very different environments surround bacteria and mitochondria, and we discuss the role of Sam35 and Sam37 in terms of the problems peculiar to mitochondrial protein substrates.
Figures
Similar articles
-
From evolution to pathogenesis: the link between β-barrel assembly machineries in the outer membrane of mitochondria and gram-negative bacteria.Int J Mol Sci. 2012;13(7):8038-8050. doi: 10.3390/ijms13078038. Epub 2012 Jun 28. Int J Mol Sci. 2012. PMID: 22942688 Free PMC article. Review.
-
Sam37 is crucial for formation of the mitochondrial TOM-SAM supercomplex, thereby promoting β-barrel biogenesis.J Cell Biol. 2015 Sep 28;210(7):1047-54. doi: 10.1083/jcb.201504119. J Cell Biol. 2015. PMID: 26416958 Free PMC article.
-
Genetic and functional interactions between the mitochondrial outer membrane proteins Tom6 and Sam37.Mol Cell Biol. 2009 Nov;29(22):5975-88. doi: 10.1128/MCB.00069-09. Epub 2009 Sep 21. Mol Cell Biol. 2009. PMID: 19797086 Free PMC article.
-
Mitochondrial sorting and assembly machinery operates by β-barrel switching.Nature. 2021 Feb;590(7844):163-169. doi: 10.1038/s41586-020-03113-7. Epub 2021 Jan 6. Nature. 2021. PMID: 33408415
-
Biogenesis of mitochondrial β-barrel membrane proteins.FEBS Open Bio. 2024 Oct;14(10):1595-1609. doi: 10.1002/2211-5463.13905. Epub 2024 Sep 29. FEBS Open Bio. 2024. PMID: 39343721 Free PMC article. Review.
Cited by
-
From evolution to pathogenesis: the link between β-barrel assembly machineries in the outer membrane of mitochondria and gram-negative bacteria.Int J Mol Sci. 2012;13(7):8038-8050. doi: 10.3390/ijms13078038. Epub 2012 Jun 28. Int J Mol Sci. 2012. PMID: 22942688 Free PMC article. Review.
-
Ancestral and derived protein import pathways in the mitochondrion of Reclinomonas americana.Mol Biol Evol. 2011 May;28(5):1581-91. doi: 10.1093/molbev/msq305. Epub 2010 Nov 15. Mol Biol Evol. 2011. PMID: 21081480 Free PMC article.
-
In vivo roles of BamA, BamB and BamD in the biogenesis of BamA, a core protein of the β-barrel assembly machine of Escherichia coli.J Mol Biol. 2015 Mar 13;427(5):1061-74. doi: 10.1016/j.jmb.2014.04.021. Epub 2014 May 2. J Mol Biol. 2015. PMID: 24792419 Free PMC article.
-
A Biochemical and Structural Understanding of TOM Complex Interactions and Implications for Human Health and Disease.Cells. 2021 May 11;10(5):1164. doi: 10.3390/cells10051164. Cells. 2021. PMID: 34064787 Free PMC article. Review.
-
Cytosolic Hsp70 and Hsp40 chaperones enable the biogenesis of mitochondrial β-barrel proteins.J Cell Biol. 2018 Sep 3;217(9):3091-3108. doi: 10.1083/jcb.201712029. Epub 2018 Jun 21. J Cell Biol. 2018. PMID: 29930205 Free PMC article.
References
-
- Armstrong L. C., Komiya T., Bergman B. E., Mihara K., Bornstein P. Metaxin is a component of a preprotein import complex in the outer membrane of the mammalian mitochondrion. J. Biol. Chem. 1997;272:6510–6518. - PubMed
-
- Buchanan S. K. Beta-barrel proteins from bacterial outer membranes: structure, function and refolding. Curr. Opin. Struct. Biol. 1999;9:455–461. - PubMed
-
- Daum G., Bohni P. C., Schatz G. Import of proteins into mitochondria. Cytochrome b2 and cytochrome c peroxidase are located in the intermembrane space of yeast mitochondria. J. Biol. Chem. 1982;257:13028–13033. - PubMed
-
- Dolezal P., Likic V., Tachezy J., Lithgow T. Evolution of the molecular machines for protein import into mitochondria. Science. 2006;313:314–318. - PubMed
-
- Gabriel K., Buchanan S. K., Lithgow T. The alpha and the beta: protein translocation across mitochondrial and plastid outer membranes. Trends Biochem. Sci. 2001;26:36–40. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
