

Dynamic association of the fragile X mental retardation protein as a messenger ribonucleoprotein between microtubules and polyribosomes
- PMID: 17978095
- PMCID: PMC2174202
- DOI: 10.1091/mbc.e07-06-0583
Dynamic association of the fragile X mental retardation protein as a messenger ribonucleoprotein between microtubules and polyribosomes
Abstract
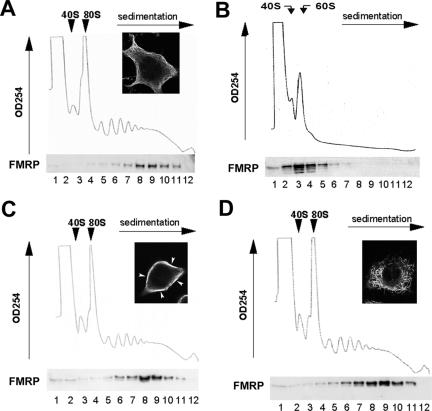
The fragile X mental retardation protein (FMRP) is a selective RNA-binding protein that regulates translation and plays essential roles in synaptic function. FMRP is bound to specific mRNA ligands, actively transported into neuronal processes in a microtubule-dependent manner, and associated with polyribosomes engaged in translation elongation. However, the biochemical relationship between FMRP-microtubule association and FMRP-polyribosome association remains elusive. Here, we report that although the majority of FMRP is incorporated into elongating polyribosomes in the soluble cytoplasm, microtubule-associated FMRP is predominantly retained in translationally dormant, polyribosome-free messenger ribonucleoprotein (mRNP) complexes. Interestingly, FMRP-microtubule association is increased when mRNPs are dynamically released from polyribosomes as a result of inhibiting translation initiation. Furthermore, the I304N mutant FMRP that fails to be incorporated into polyribosomes is associated with microtubules in mRNP particles and transported into neuronal dendrites in a microtubule-dependent, 3,5-dihydroxyphenylglycine-stimulated manner with similar kinetics to that of wild-type FMRP. Hence, polyribosome-free FMRP-mRNP complexes travel on microtubules and wait for activity-dependent translational derepression at the site of function. The dual participation of FMRP in dormant mRNPs and polyribosomes suggests distinct roles of FMRP in dendritic transport and translational regulation, two distinct phases that control local protein production to accommodate synaptic plasticity.
Figures






References
-
- Antar L. N., Bassell G. J. Sunrise at the synapse: the FMRP mRNP shaping the synaptic interface. Neuron. 2003;37:555–558. - PubMed
-
- Antar L. N., Dictenberg J. B., Plociniak M., Afroz R., Bassell G. J. Localization of FMRP-associated mRNA granules and requirement of microtubules for activity-dependent trafficking in hippocampal neurons. Genes Brain Behav. 2005;4:350–359. - PubMed
-
- Bagni C., Greenough W. T. From mRNP trafficking to spine dysmorphogenesis: the roots of fragile X syndrome. Nat. Rev. Neurosci. 2005;6:376–387. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
