

Human capping enzyme promotes formation of transcriptional R loops in vitro
- PMID: 17978174
- PMCID: PMC2077024
- DOI: 10.1073/pnas.0708866104
Human capping enzyme promotes formation of transcriptional R loops in vitro
Abstract
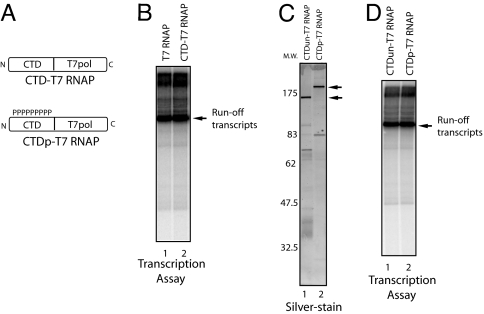
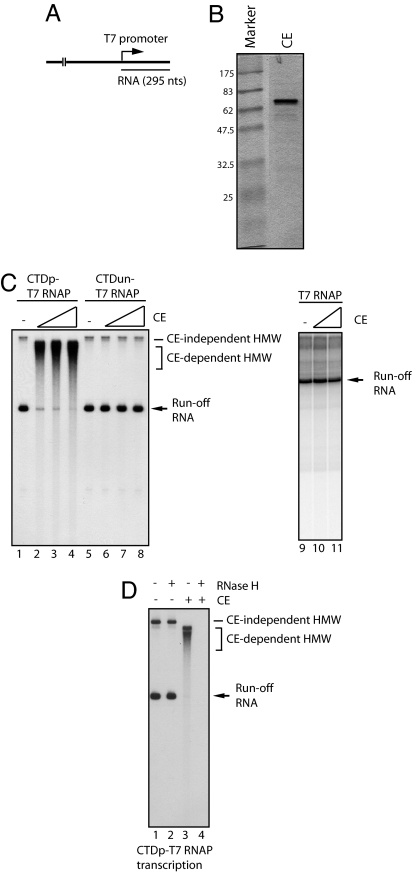
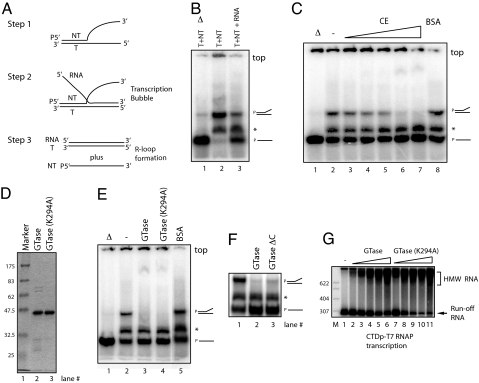
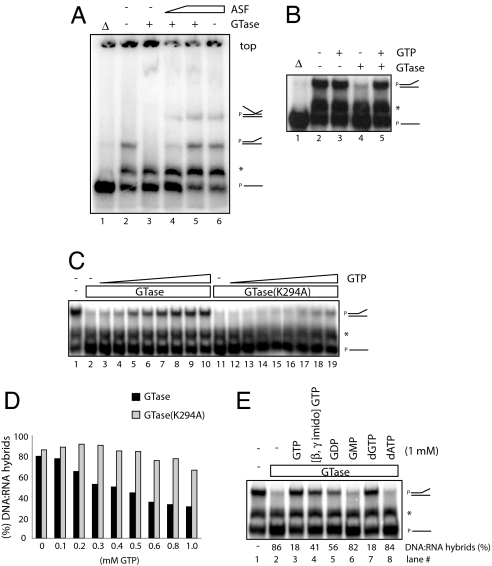
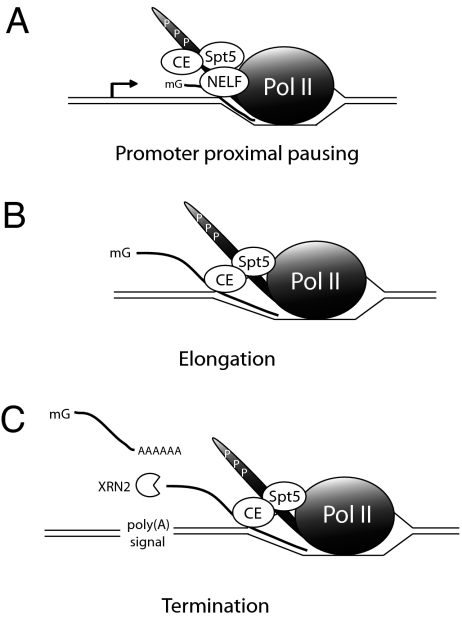
Cap formation is the first step of pre-mRNA processing in eukaryotic cells. Immediately after transcription initiation, capping enzyme (CE) is recruited to RNA polymerase II (Pol II) by the phosphorylated carboxyl-terminal domain of the Pol II largest subunit (CTD), allowing cotranscriptional capping of the nascent pre-mRNA. Recent studies have indicated that CE affects transcription elongation and have suggested a checkpoint model in which cotranscriptional capping is a necessary step for the early phase of transcription. To investigate further the role of the CTD in linking transcription and processing, we generated a fusion protein of the mouse CTD with T7 RNA polymerase (CTD-T7 RNAP). Unexpectedly, in vitro transcription assays with CTD-T7 RNAP showed that CE promotes formation of DNA.RNA hybrids or R loops. Significantly, phosphorylation of the CTD was required for CE-dependent R-loop formation (RLF), consistent with a critical role for the CTD in CE recruitment to the transcription complex. The guanylyltransferase domain was necessary and sufficient for RLF, but catalytic activity was not required. In vitro assays with appropriate synthetic substrates indicate that CE can promote RLF independent of transcription. ASF/SF2, a splicing factor known to prevent RLF, and GTP, which affects CE conformation, antagonized CE-dependent RLF. Our findings suggest that CE can play a direct role in transcription by modulating displacement of nascent RNA during transcription.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
The Carboxyl-terminal Domain of RNA Polymerase II Is Not Sufficient to Enhance the Efficiency of Pre-mRNA Capping or Splicing in the Context of a Different Polymerase.J Biol Chem. 2009 Mar 27;284(13):8692-702. doi: 10.1074/jbc.M806919200. Epub 2009 Jan 28. J Biol Chem. 2009. PMID: 19176527 Free PMC article.
-
A novel RNA pol II CTD interaction site on the mRNA capping enzyme is essential for its allosteric activation.Nucleic Acids Res. 2021 Apr 6;49(6):3109-3126. doi: 10.1093/nar/gkab130. Nucleic Acids Res. 2021. PMID: 33684220 Free PMC article.
-
Molecular Basis of Transcription-Coupled Pre-mRNA Capping.Mol Cell. 2015 Jun 18;58(6):1079-89. doi: 10.1016/j.molcel.2015.04.004. Epub 2015 May 7. Mol Cell. 2015. PMID: 25959396
-
Phosphorylation of the C-terminal domain of RNA polymerase II.Biochim Biophys Acta. 1995 Apr 4;1261(2):171-82. doi: 10.1016/0167-4781(94)00233-s. Biochim Biophys Acta. 1995. PMID: 7711060 Review.
-
The CTD role in cotranscriptional RNA processing and surveillance.FEBS Lett. 2008 Jun 18;582(14):1971-6. doi: 10.1016/j.febslet.2008.04.019. Epub 2008 Apr 22. FEBS Lett. 2008. PMID: 18435923 Review.
Cited by
-
RNA-DNA strand exchange by the Drosophila Polycomb complex PRC2.Nat Commun. 2020 Apr 14;11(1):1781. doi: 10.1038/s41467-020-15609-x. Nat Commun. 2020. PMID: 32286294 Free PMC article.
-
G clustering is important for the initiation of transcription-induced R-loops in vitro, whereas high G density without clustering is sufficient thereafter.Mol Cell Biol. 2009 Jun;29(11):3124-33. doi: 10.1128/MCB.00139-09. Epub 2009 Mar 23. Mol Cell Biol. 2009. PMID: 19307304 Free PMC article.
-
Mechanism of expression regulation of head-to-head overlapping protein-coding genes INO80E and HIRIP3.Commun Biol. 2025 Mar 8;8(1):391. doi: 10.1038/s42003-025-07815-4. Commun Biol. 2025. PMID: 40057624 Free PMC article.
-
Transcription induces strand-specific mutations at the 5' end of human genes.Genome Res. 2008 Aug;18(8):1216-23. doi: 10.1101/gr.076570.108. Epub 2008 May 7. Genome Res. 2008. PMID: 18463301 Free PMC article.
-
NKAPL facilitates transcription pause-release and bridges elongation to initiation during meiosis exit.Nat Commun. 2025 Jan 17;16(1):791. doi: 10.1038/s41467-024-55579-y. Nat Commun. 2025. PMID: 39824811 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
