

SUMO-specific protease 1 is essential for stabilization of HIF1alpha during hypoxia
- PMID: 17981124
- PMCID: PMC2128732
- DOI: 10.1016/j.cell.2007.08.045
SUMO-specific protease 1 is essential for stabilization of HIF1alpha during hypoxia
Abstract
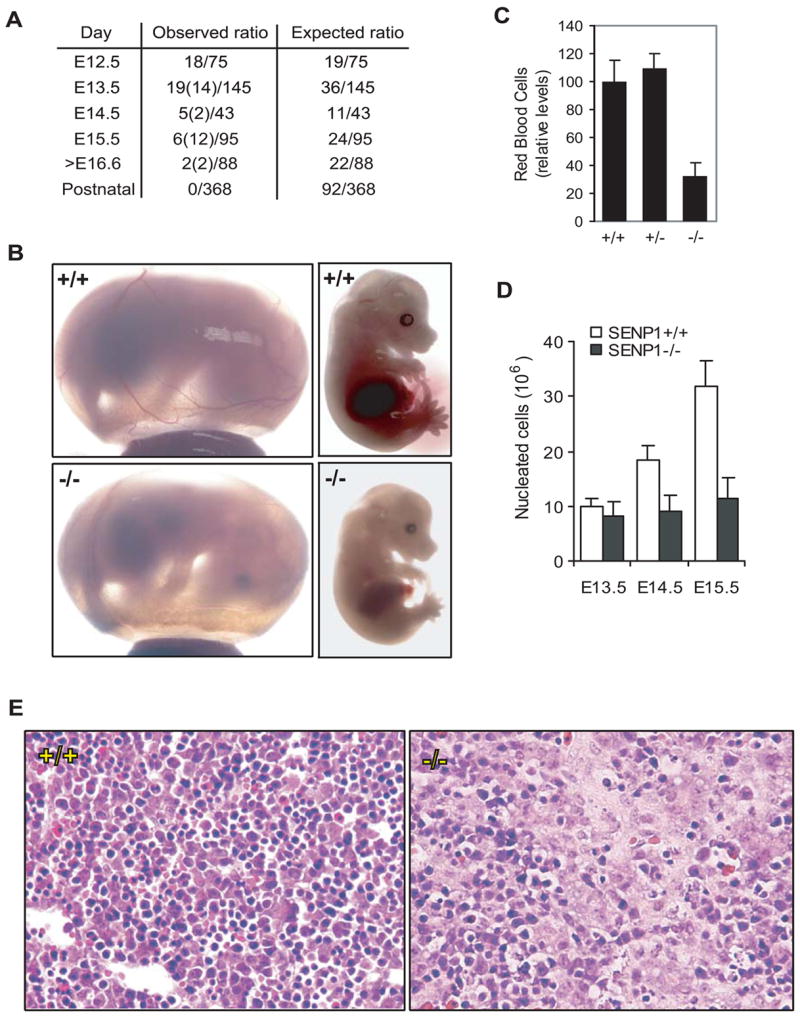
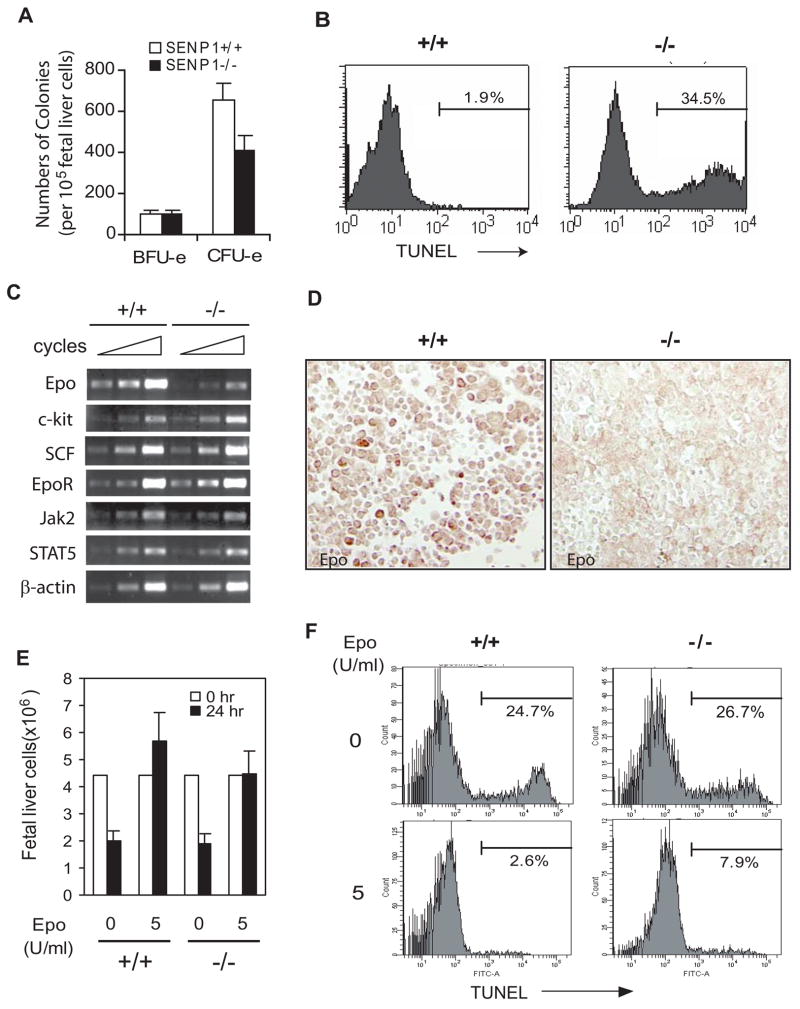
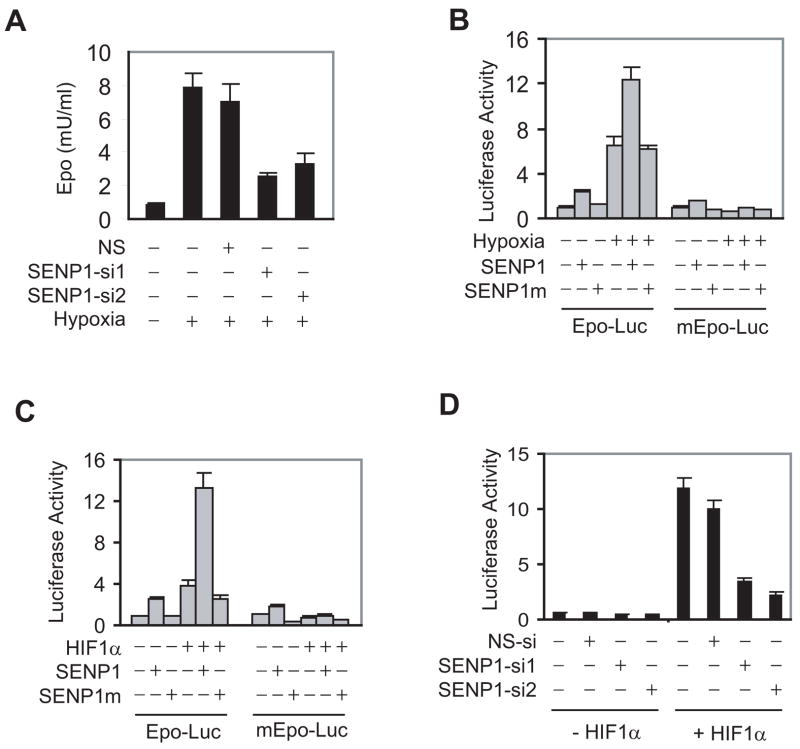
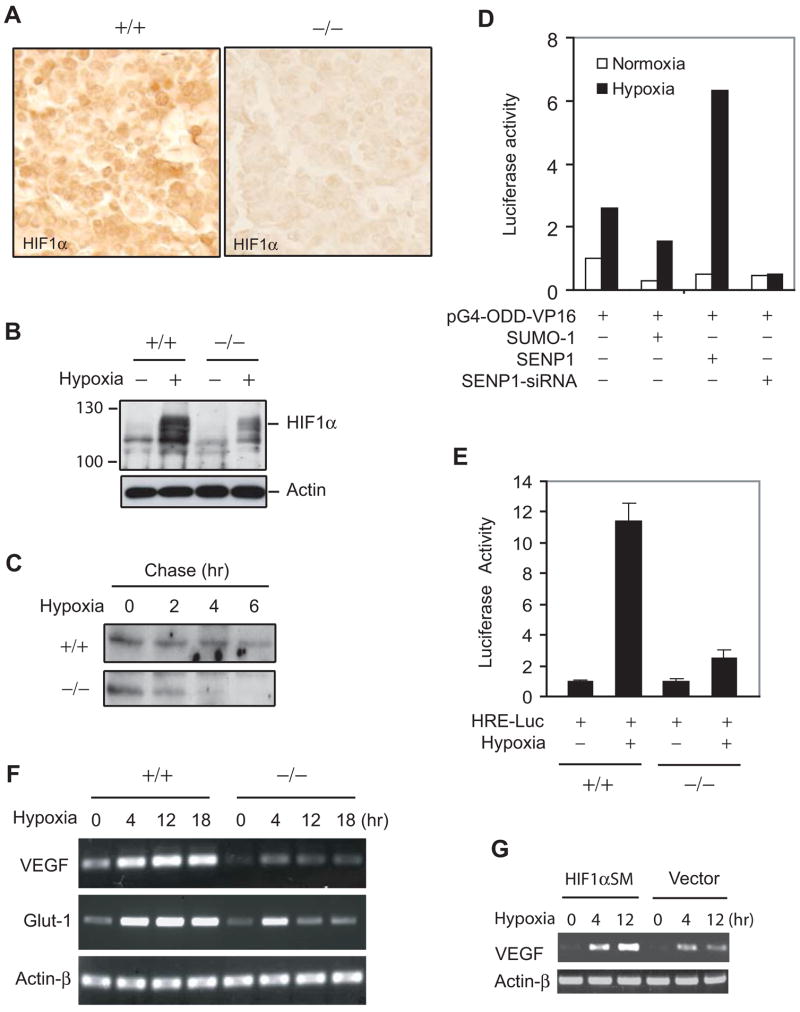
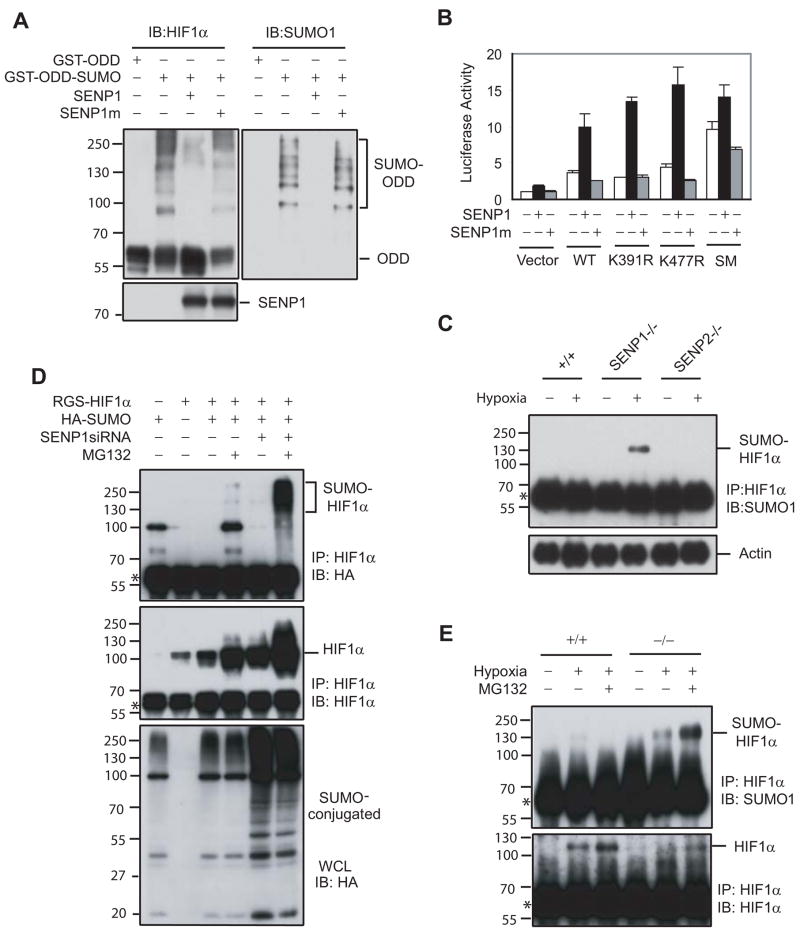
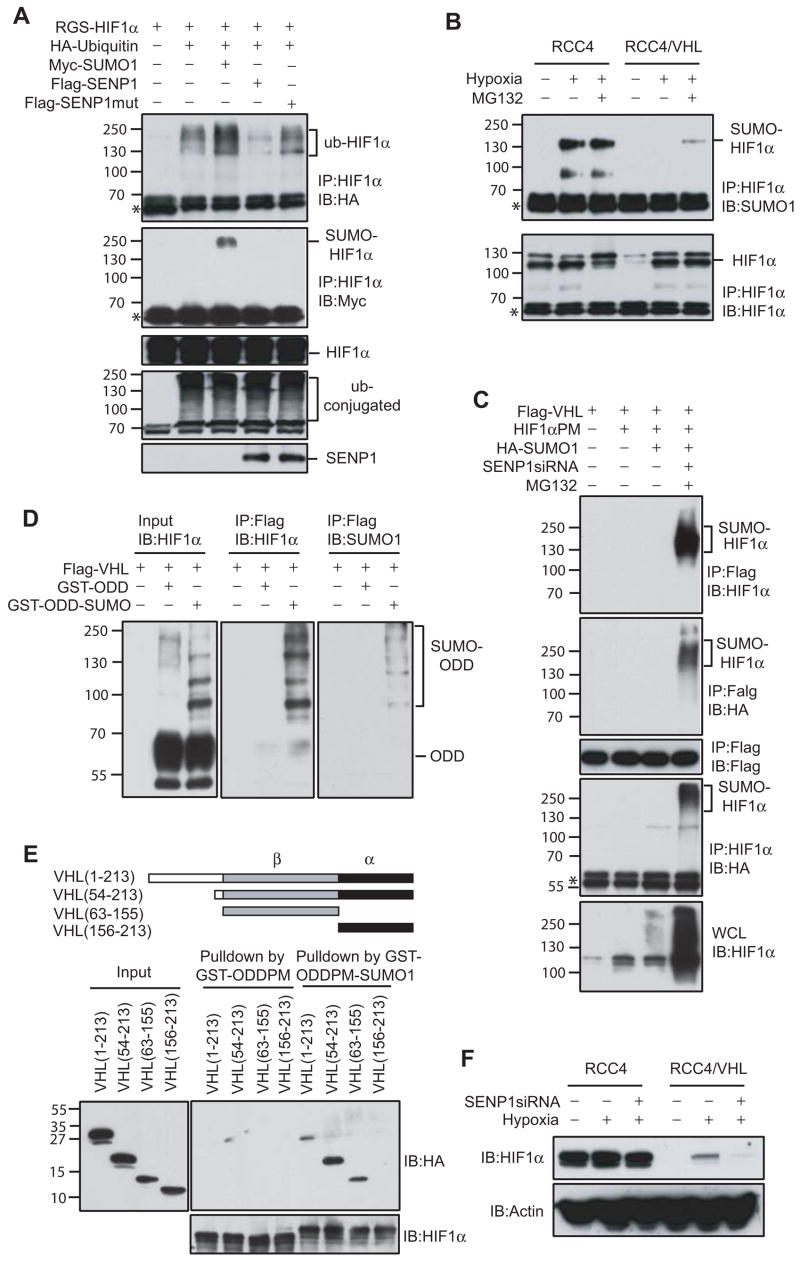
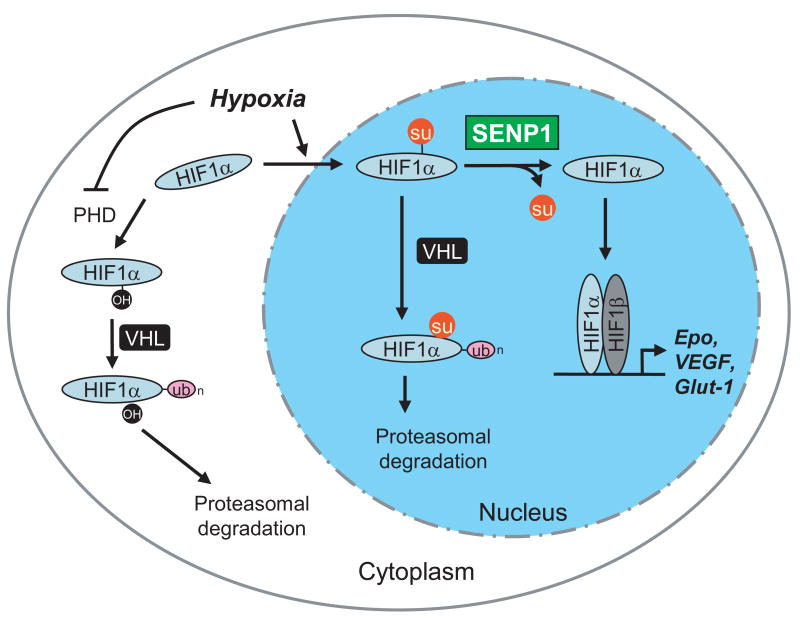
SUMOylation is a dynamic process, catalyzed by SUMO-specific ligases and reversed by Sentrin/SUMO-specific proteases (SENPs). The physiologic consequences of SUMOylation and deSUMOylation are not fully understood. Here we investigate the phenotypes of mice lacking SENP1 and find that SENP1(-/-) embryos show severe fetal anemia stemming from deficient erythropoietin (Epo) production and die midgestation. We determine that SENP1 controls Epo production by regulating the stability of hypoxia-inducible factor 1alpha (HIF1alpha) during hypoxia. Hypoxia induces SUMOylation of HIF1alpha, which promotes its binding to a ubiquitin ligase, von Hippel-Lindau (VHL) protein, through a proline hydroxylation-independent mechanism, leading to its ubiquitination and degradation. In SENP1(-/-) MEFs, hypoxia-induced transcription of HIF1alpha-dependent genes such as vascular endothelial growth factor (VEGF) and glucose transporter 1 (Glut-1) is markedly reduced. These results show that SENP1 plays a key role in the regulation of the hypoxic response through regulation of HIF1alpha stability and that SUMOylation can serve as a direct signal for ubiquitin-dependent degradation.
Figures
Comment in
-
SUMO teams up with ubiquitin to manage hypoxia.Cell. 2007 Nov 2;131(3):446-7. doi: 10.1016/j.cell.2007.10.026. Cell. 2007. PMID: 17981111
References
-
- Bae SH, Jeong JW, Park JA, Kim SH, Bae MK, Choi SJ, Kim KW. Sumoylation increases HIF-1alpha stability and its transcriptional activity. Biochem Biophys Res Commun. 2004;324:394–400. - PubMed
-
- Brahimi-Horn C, Mazure N, Pouyssegur J. Signalling via the hypoxia-inducible factor-1alpha requires multiple posttranslational modifications. Cell Signal. 2005;17:1–9. - PubMed
-
- Bruick RK, McKnight SL. A conserved family of prolyl-4-hydroxylases that modify HIF. Science. 2001;294:1337–1340. - PubMed
-
- Cheng J, Perkins ND, Yeh ET. Differential Regulation of c-Jun-dependent Transcription by SUMO-specific Proteases. J Biol Chem. 2005;280:14492–14498. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
