

The role of the mitochondrial glycine cleavage complex in the metabolism and virulence of the protozoan parasite Leishmania major
- PMID: 17981801
- PMCID: PMC2963101
- DOI: 10.1074/jbc.M708014200
The role of the mitochondrial glycine cleavage complex in the metabolism and virulence of the protozoan parasite Leishmania major
Abstract
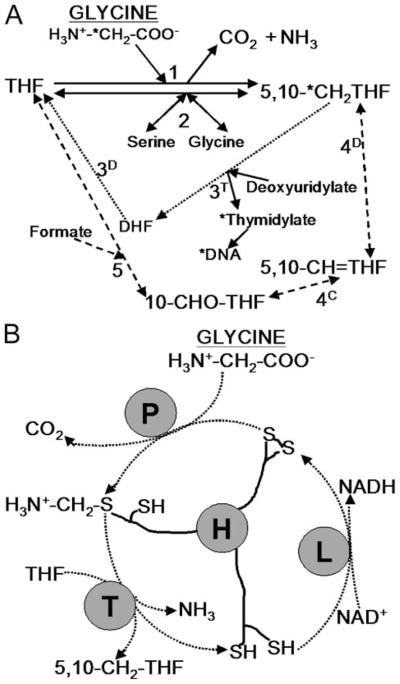
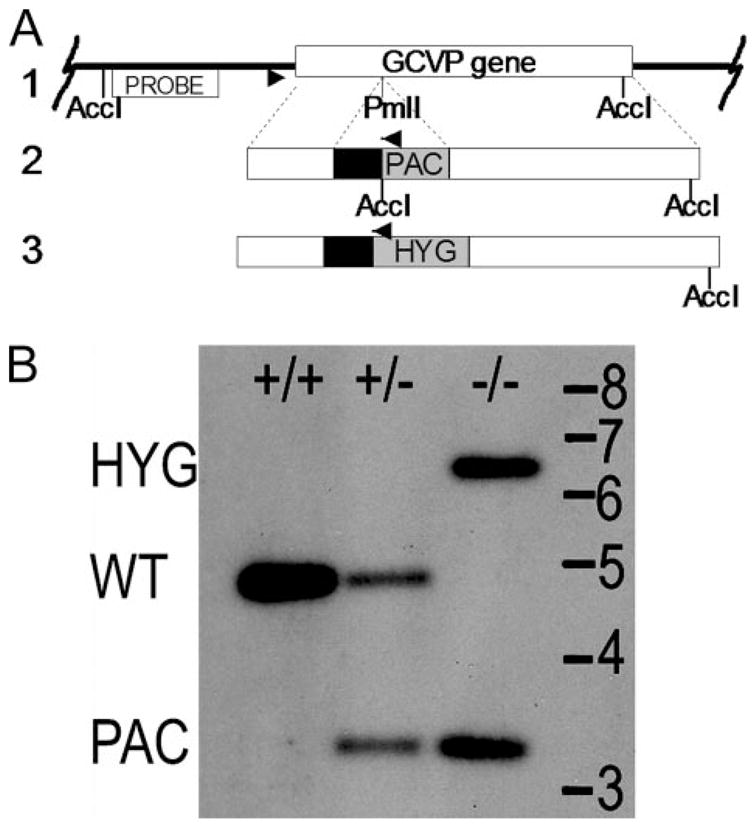
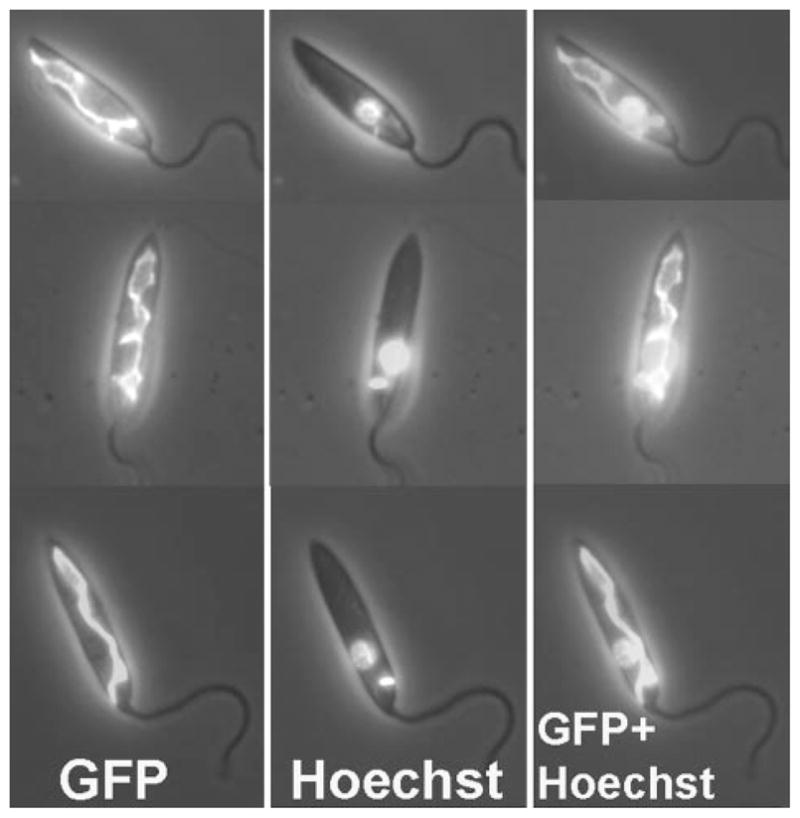
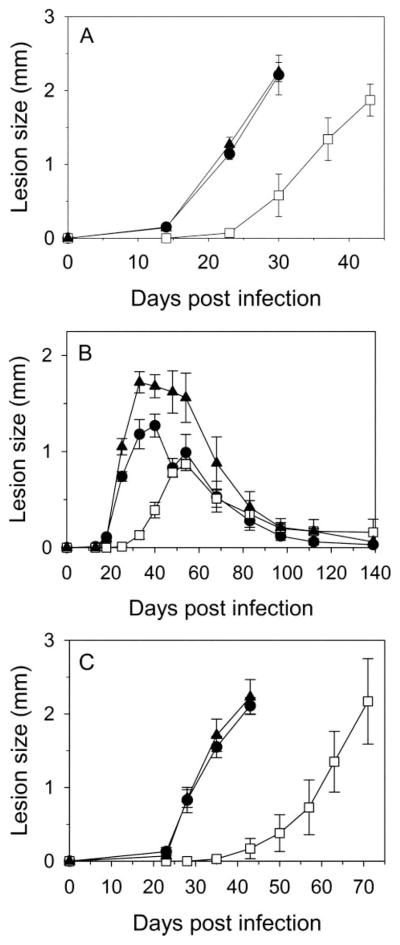
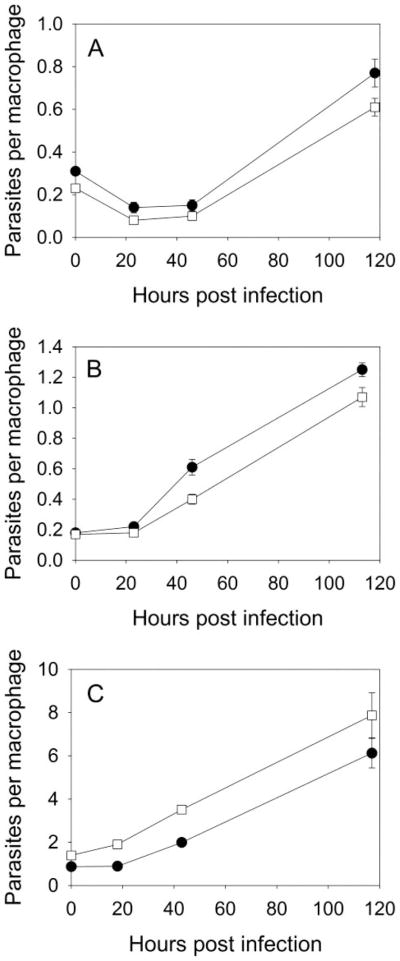
For the human pathogen Leishmania major, a key metabolic function is the synthesis of thymidylate, which requires 5,10-methylenetetrahydrofolate (5,10-CH(2)-THF). 5,10-CH(2)-THF can be synthesized from glycine by the mitochondrial glycine cleavage complex (GCC). Bioinformatic analysis revealed the four subunits of the GCC in the L. major genome, and the role of the GCC in parasite metabolism and virulence was assessed through studies of the P subunit (glycine decarboxylase (GCVP)). First, a tagged GCVP protein was expressed and localized to the parasite mitochondrion. Second, a gcvP(-) mutant was generated and shown to lack significant GCC activity using an indirect in vivo assay after incorporation of label from [2-(14)C]glycine into DNA. The gcvP(-) mutant grew poorly in the presence of excess glycine or minimal serine; these studies also established that L. major promastigotes require serine for optimal growth. Although gcvP(-) promastigotes and amastigotes showed normal virulence in macrophage infections in vitro, both forms of the parasite showed substantially delayed replication and lesion pathology in infections of both genetically susceptible or resistant mice. These data suggest that, as the physiology of the infection site changes during the course of infection, so do the metabolic constraints on parasite replication. This conclusion has great significance to the interpretation of metabolic requirements for virulence. Last, these studies call attention in trypanosomatid protozoa to the key metabolic intermediate 5,10-CH(2)-THF, situated at the junction of serine, glycine, and thymidylate metabolism. Notably, genome-based predictions suggest the related parasite Trypanosoma brucei is totally dependent on the GCC for 5,10-CH(2)-THF synthesis.
Figures
References
-
- Herwaldt BL. Lancet. 1999;354:1191–1199. - PubMed
-
- Lira R, Sundar S, Makharia A, Kenney R, Gam A, Saraiva E, Sacks D. J Infect Dis. 1999;180:564–567. - PubMed
-
- Berman J, Bryceson AD, Croft S, Engel J, Gutteridge W, Karbwang J, Sindermann H, Soto J, Sundar S, Urbina JA. Trans R Soc Trop Med Hyg. 2006;100(Suppl 1):41–44. - PubMed
-
- Garrett CE, Coderre JA, Meek TD, Garvey EP, Claman DM, Beverley SM, Santi DV. Mol Biochem Parasitol. 1984;11:257–265. - PubMed
-
- Stechmann A, Cavalier-Smith T. Science. 2002;297:89–91. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
