

Characterization of the G-quadruplexes in the duplex nuclease hypersensitive element of the PDGF-A promoter and modulation of PDGF-A promoter activity by TMPyP4
- PMID: 17984069
- PMCID: PMC2190695
- DOI: 10.1093/nar/gkm538
Characterization of the G-quadruplexes in the duplex nuclease hypersensitive element of the PDGF-A promoter and modulation of PDGF-A promoter activity by TMPyP4
Abstract
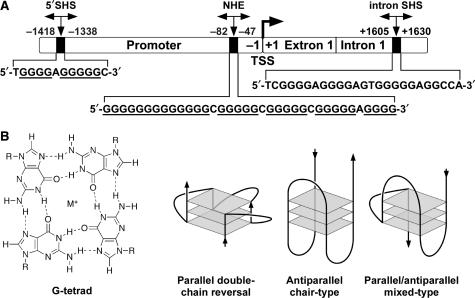
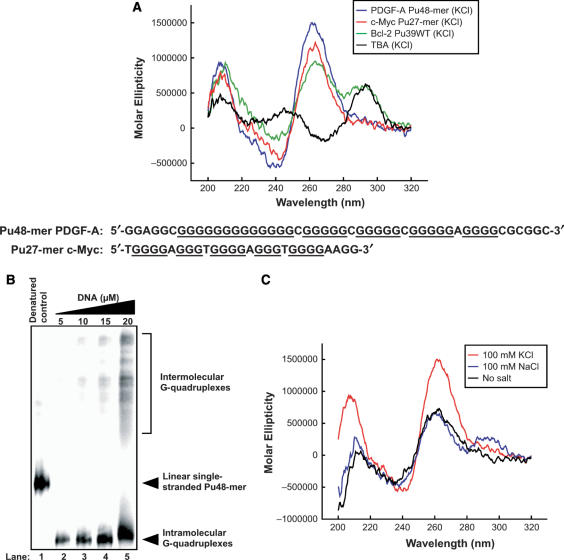
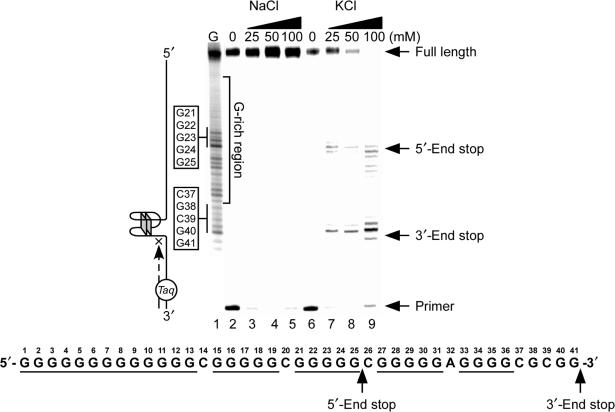
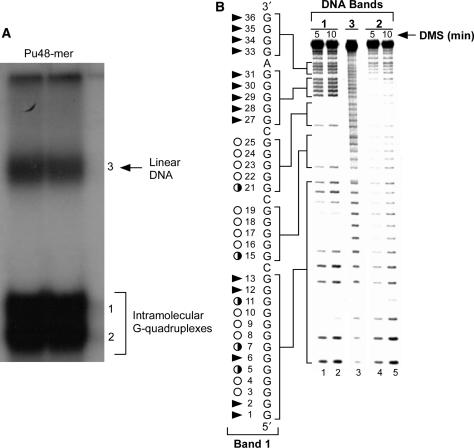
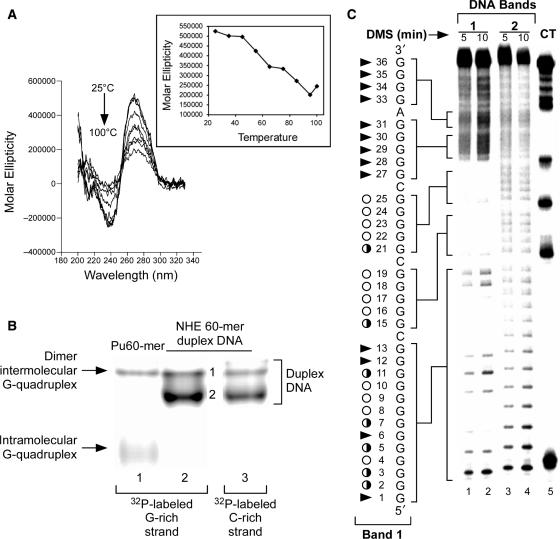
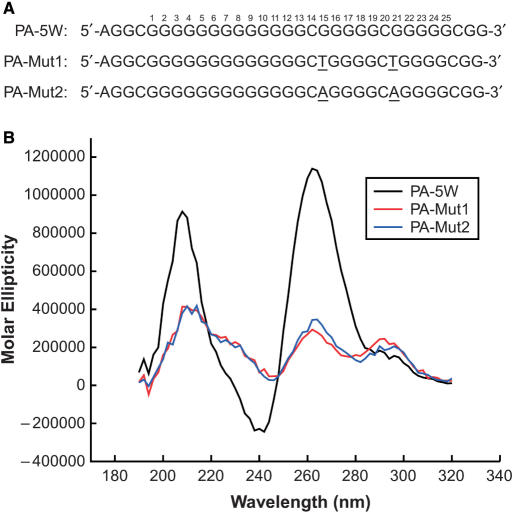
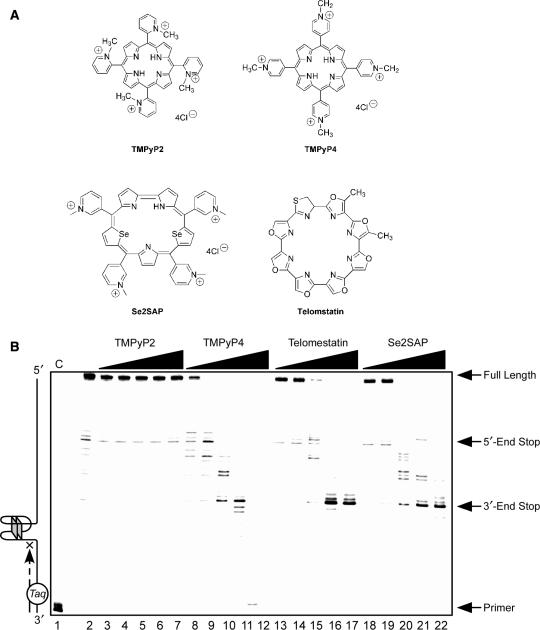
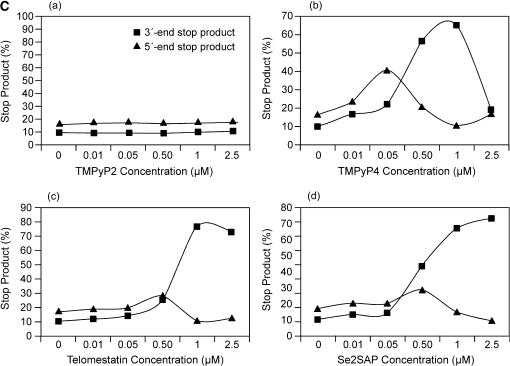
The proximal 5'-flanking region of the human platelet-derived growth factor A (PDGF-A) promoter contains one nuclease hypersensitive element (NHE) that is critical for PDGF-A gene transcription. On the basis of circular dichroism (CD) and electrophoretic mobility shift assay (EMSA), we have shown that the guanine-rich (G-rich) strand of the DNA in this region can form stable intramolecular parallel G-quadruplexes under physiological conditions. A Taq polymerase stop assay has shown that the G-rich strand of the NHE can form two major G-quadruplex structures, which are in dynamic equilibrium and differentially stabilized by three G-quadruplex-interactive drugs. One major parallel G-quadruplex structure of the G-rich strand DNA of NHE was identified by CD and dimethyl sulfate (DMS) footprinting. Surprisingly, CD spectroscopy shows a stable parallel G-quadruplex structure formed within the duplex DNA of the NHE at temperatures up to 100 degrees C. This structure has been characterized by DMS footprinting in the double-stranded DNA of the NHE. In transfection experiments, 10 microM TMPyP4 reduced the activity of the basal promoter of PDGF-A approximately 40%, relative to the control. On the basis of these results, we have established that ligand-mediated stabilization of G-quadruplex structures within the PDGF-A NHE can silence PDGF-A expression.
Figures
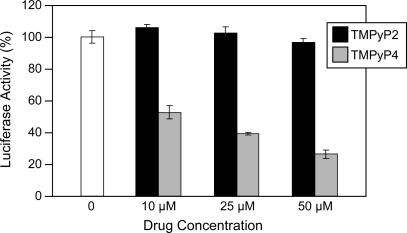
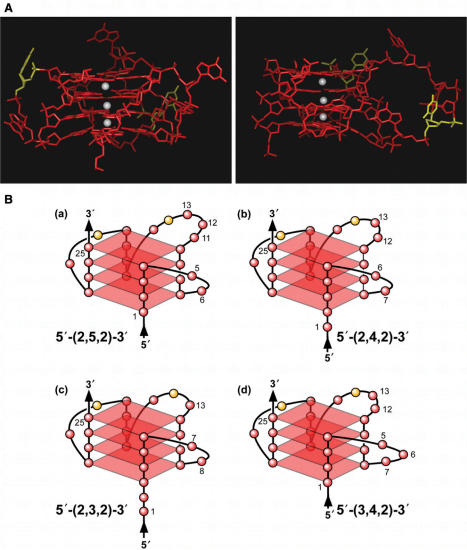
References
-
- Heldin CH, Westermark B. Mechanism of action and in vivo role of platelet-derived growth factor. Physiol. Rev. 1999;79:1283–1316. - PubMed
-
- Heldin CH, Eriksson U, Ostman A. New members of the platelet-derived growth factor family of mitogens. Arch. Biochem. Biophys. 2002;398:284–290. - PubMed
-
- Alvarez RH, Kantarjian HM, Cortes JE. Biology of platelet-derived growth factor and its involvement in disease. Mayo Clin. Proc. 2006;81:1241–1257. - PubMed
-
- Betsholtz C. Biology of platelet-derived growth factors in development. Birth Defects Res. C Embryo. Today. 2003;69:272–285. - PubMed
-
- Ostman A, Heldin CH. Involvement of platelet-derived growth factor in disease: development of specific antagonists. Adv. Cancer Res. 2001;80:1–38. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
