

Nuclear receptor interaction protein, a coactivator of androgen receptors (AR), is regulated by AR and Sp1 to feed forward and activate its own gene expression through AR protein stability
- PMID: 17984071
- PMCID: PMC2248731
- DOI: 10.1093/nar/gkm942
Nuclear receptor interaction protein, a coactivator of androgen receptors (AR), is regulated by AR and Sp1 to feed forward and activate its own gene expression through AR protein stability
Abstract
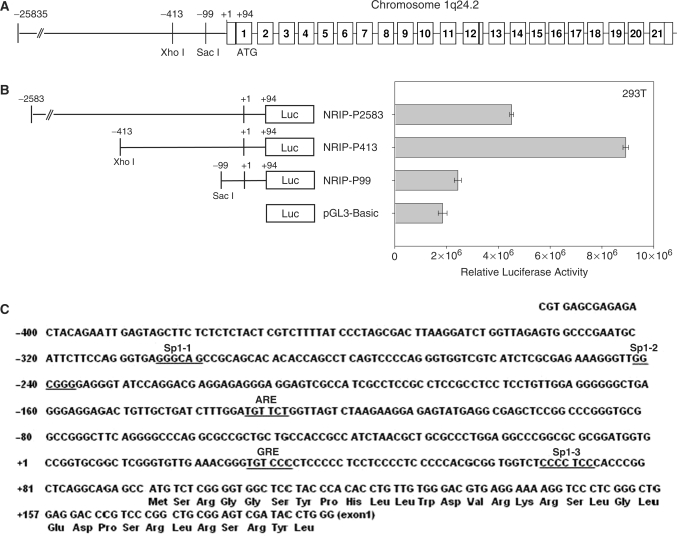
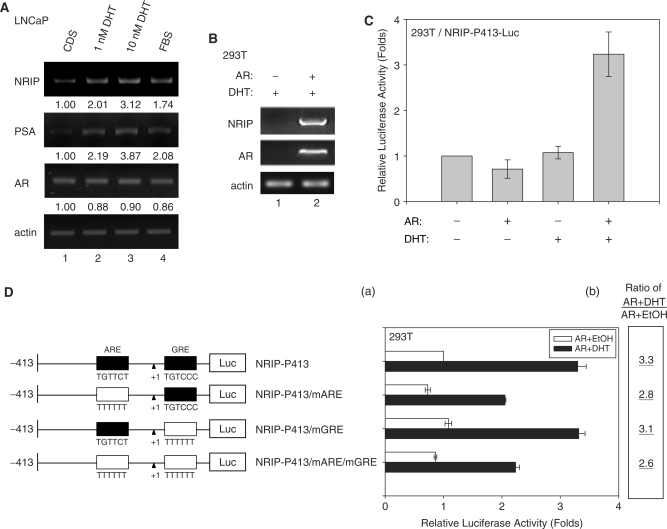
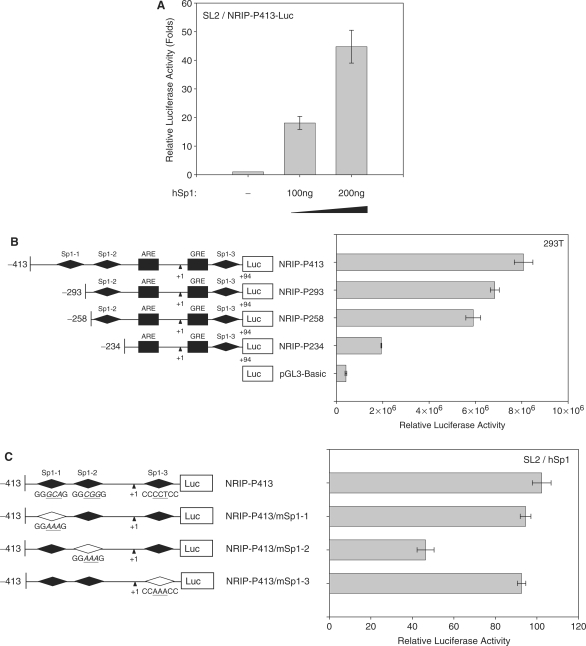
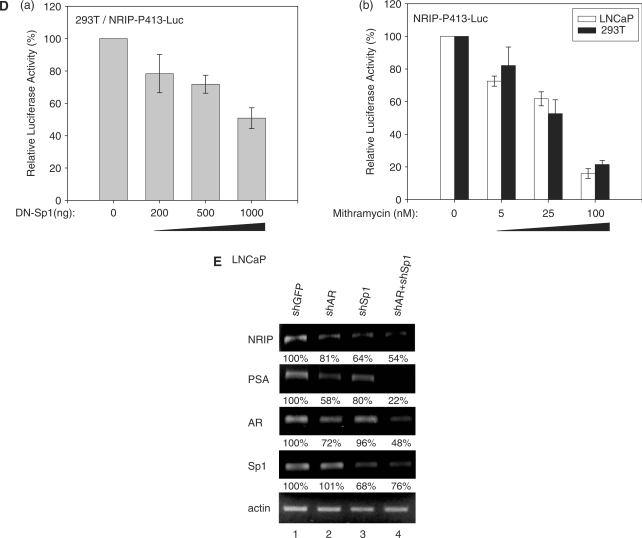
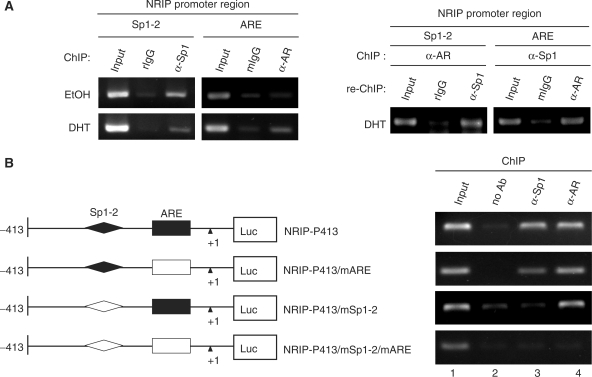
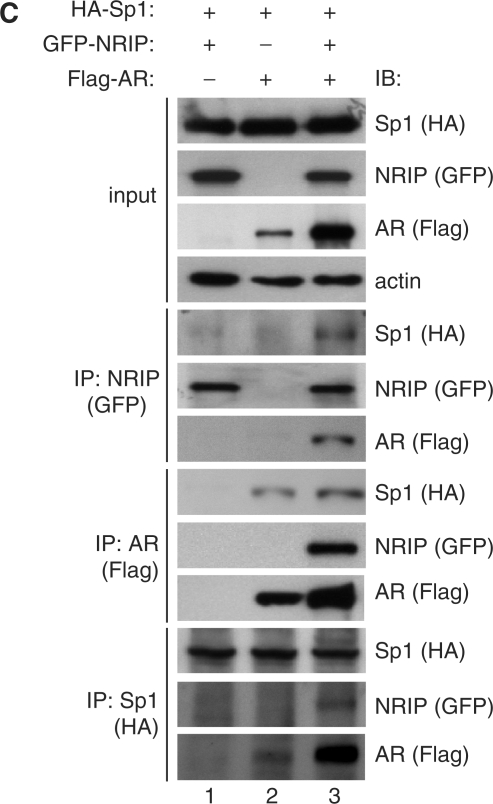
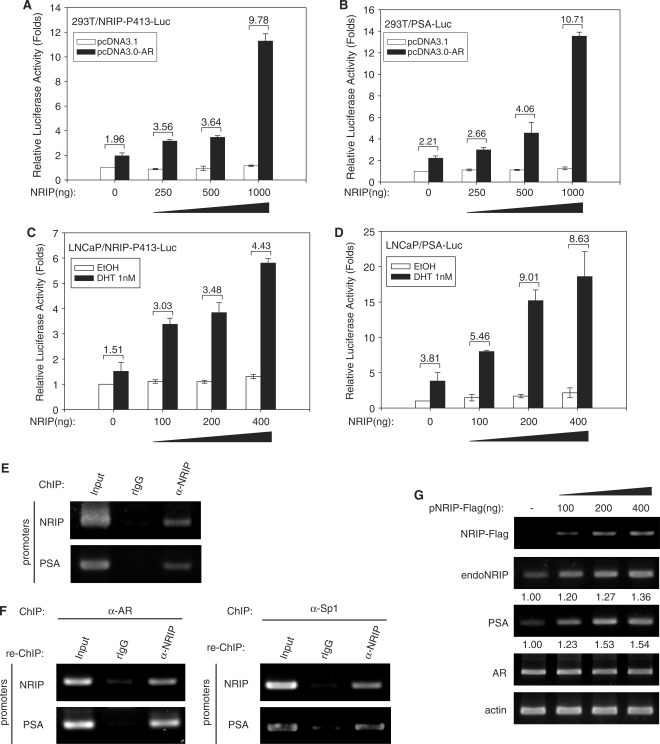
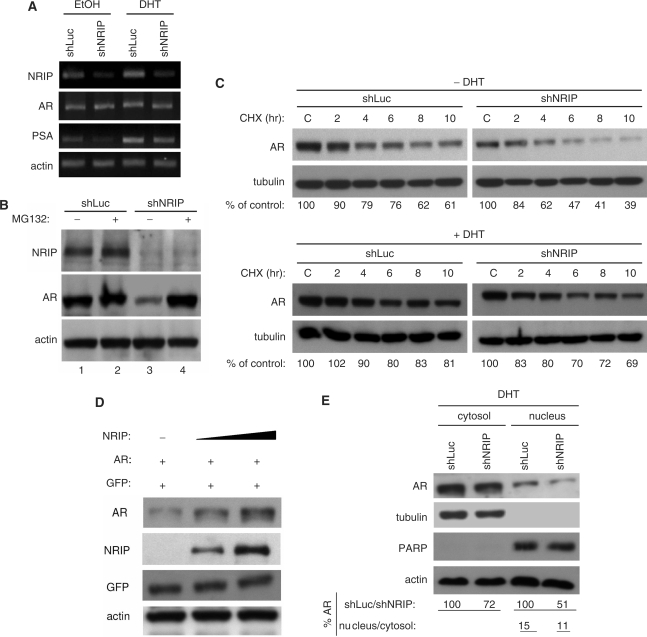
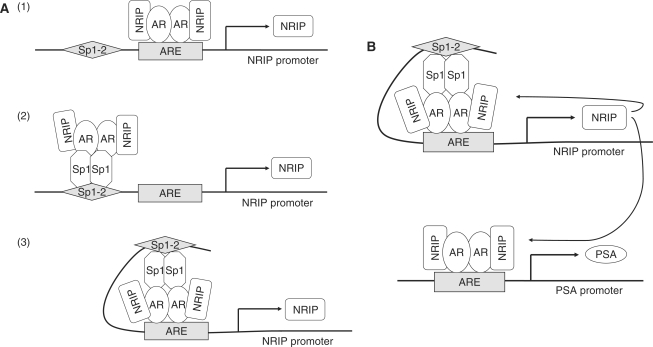
Previously, we found a novel gene, nuclear receptor interaction protein (NRIP), a transcription cofactor that can enhance an AR-driven PSA promoter activity in a ligand-dependent manner in prostate cancer cells. Here, we investigated NRIP regulation. We cloned a 413-bp fragment from the transcription initiation site of the NRIP gene that had strong promoter activity, was TATA-less and GC-rich, and, based on DNA sequences, contained one androgen response element (ARE) and three Sp1-binding sites (Sp1-1, Sp1-2, Sp1-3). Transient promoter luciferase assays, chromatin immunoprecipitation and small RNA interference analyses mapped ARE and Sp1-2-binding sites involved in NRIP promoter activation, implying that NRIP is a target gene for AR or Sp1. AR associates with the NRIP promoter through ARE and indirectly through Sp1-binding site via AR-Sp1 complex formation. Thus both ARE and Sp1-binding site within the NRIP promoter can respond to androgen induction. More intriguingly, NRIP plays a feed-forward role enhancing AR-driven NRIP promoter activity via NRIP forming a complex with AR to protect AR protein from proteasome degradation. This is the first demonstration that NRIP is a novel AR-target gene and that NRIP expression feeds forward and activates its own expression through AR protein stability.
Figures
References
-
- Black BE, Paschal BM. Intranuclear organization and function of the androgen receptor. Trends Endocrinol. Metab. 2004;15:411–417. - PubMed
-
- Burnstein KL. Regulation of androgen receptor levels: implications for prostate cancer progression and therapy. J. Cell. Biochem. 2005;95:657–669. - PubMed
-
- Heinlein CA, Chang C. Androgen receptor (AR) coregulators: an overview. Endocr. Rev. 2002;23:175–200. - PubMed
-
- Reynolds MA, Kastury K, Groskopf J, Schalken JA, Rittenhouse H. Molecular markers for prostate cancer. Cancer Lett. 2007;249:5–13. - PubMed
-
- Kumar R, Wang RA, Barnes CJ. Coregulators and chromatin remodeling in transcriptional control. Mol. Carcinog. 2004;41:221–230. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
