

Saccharomyces cerevisiae Ebs1p is a putative ortholog of human Smg7 and promotes nonsense-mediated mRNA decay
- PMID: 17984081
- PMCID: PMC2190716
- DOI: 10.1093/nar/gkm912
Saccharomyces cerevisiae Ebs1p is a putative ortholog of human Smg7 and promotes nonsense-mediated mRNA decay
Abstract
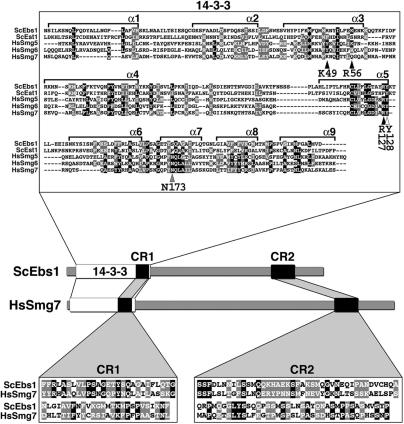
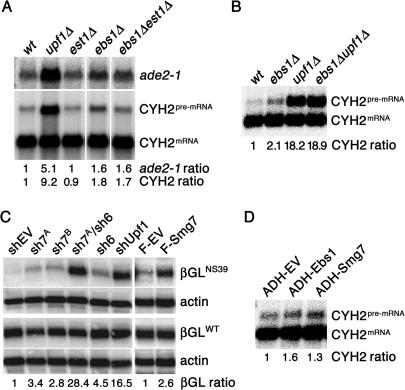
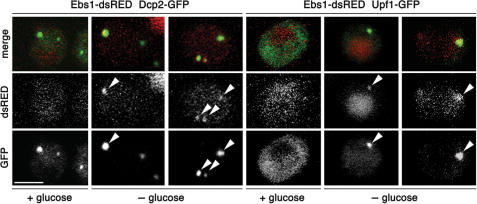
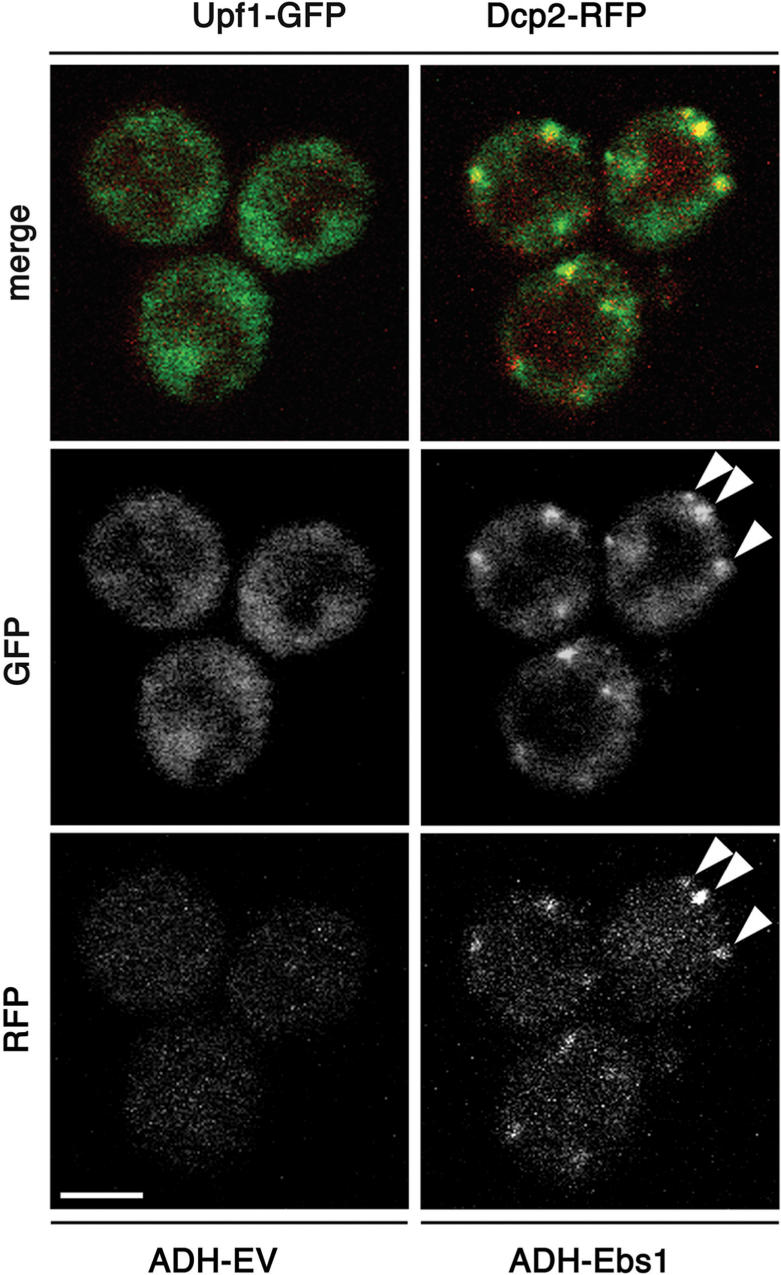
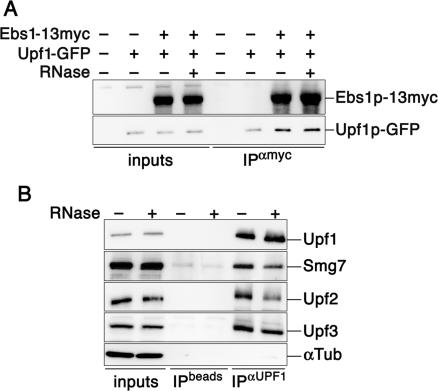
The Smg proteins Smg5, Smg6 and Smg7 are involved in nonsense-mediated RNA decay (NMD) in metazoans, but no orthologs have been found in the budding yeast Saccharomyces cerevisiae. Sequence alignments reveal that yeast Ebs1p is similar in structure to the human Smg5-7, with highest homology to Smg7. We demonstrate here that Ebs1p is involved in NMD and behaves similarly to human Smg proteins. Indeed, both loss and overexpression of Ebs1p results in stabilization of NMD targets. However, Ebs1-loss in yeast or Smg7-depletion in human cells only partially disrupts NMD and in the latter, Smg7-depletion is partially compensated for by Smg6. Ebs1p physically interacts with the NMD helicase Upf1p and overexpressed Ebs1p leads to recruitment of Upf1p into cytoplasmic P-bodies. Furthermore, Ebs1p localizes to P-bodies upon glucose starvation along with Upf1p. Overall our findings suggest that NMD is more conserved in evolution than previously thought, and that at least one of the Smg5-7 proteins is conserved in budding yeast.
Figures
References
-
- Maquat LE. Nonsense-mediated mRNA decay: splicing, translation and mRNP dynamics. Nat. Rev. Mol. Cell. Biol. 2004;5:89–99. - PubMed
-
- Azzalin CM, Lingner J. The human RNA surveillance factor UPF1 is required for S phase progression and genome stability. Curr. Biol. 2006;16:433–439. - PubMed
-
- Leeds P, Peltz SW, Jacobson A, Culbertson MR. The product of the yeast UPF1 gene is required for rapid turnover of mRNAs containing a premature translational termination codon. Genes Dev. 1991;5:2303–2314. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
