

MicroRNA399 is a long-distance signal for the regulation of plant phosphate homeostasis
- PMID: 17988220
- PMCID: PMC2268993
- DOI: 10.1111/j.1365-313X.2007.03363.x
MicroRNA399 is a long-distance signal for the regulation of plant phosphate homeostasis
Abstract
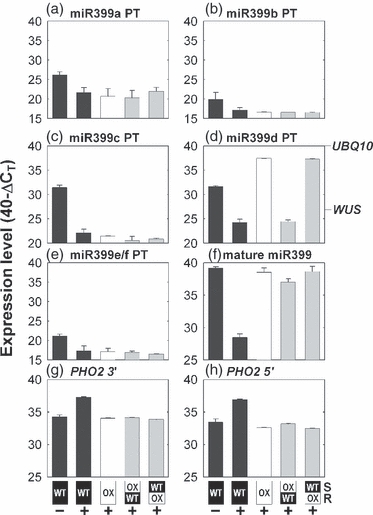
The presence of microRNA species in plant phloem sap suggests potential signaling roles by long-distance regulation of gene expression. Proof for such a role for a phloem-mobile microRNA is lacking. Here we show that phosphate (Pi) starvation-induced microRNA399 (miR399) is present in the phloem sap of two diverse plant species, rapeseed and pumpkin, and levels are strongly and specifically increased in phloem sap during Pi deprivation. By performing micro-grafting experiments using Arabidopsis, we further show that chimeric plants constitutively over-expressing miR399 in the shoot accumulate mature miR399 species to very high levels in their wild-type roots, while corresponding primary transcripts are virtually absent in roots, demonstrating shoot-to-root transport. The chimeric plants exhibit (i) down-regulation of the miR399 target transcript (PHO2), which encodes a critical component for maintenance of Pi homeostasis, in the wild-type root, and (ii) Pi accumulation in the shoot, which is the phenotype of pho2 mutants, miR399 over-expressers or chimeric plants with a genetic knock-out of PHO2 in the root. Hence the transported miR399 molecules retain biological activity. This is a demonstration of systemic control of a biological process, i.e. maintenance of plant Pi homeostasis, by a phloem-mobile microRNA.
Figures




References
-
- Allen E, Xie Z, Gustafson AM, Carrington JC. MicroRNA-directed phasing during trans-acting siRNA biogenesis in plants. Cell. 2005;121:207–221. - PubMed
-
- Bariola PA, Howard CJ, Taylor CB, Verburg MT, Jaglan VD, Green PJ. The Arabidopsis ribonuclease gene RNS1 is tightly controlled in response to phosphate limitation. Plant J. 1994;6:673–685. - PubMed
-
- Bates R, Lynch JP. Root hairs confer a competitive advantage under low phosphate availability. Plant Soil. 2001;236:243–250.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
