

The limits of subfunctionalization
- PMID: 17988397
- PMCID: PMC2213666
- DOI: 10.1186/1471-2148-7-213
The limits of subfunctionalization
Abstract
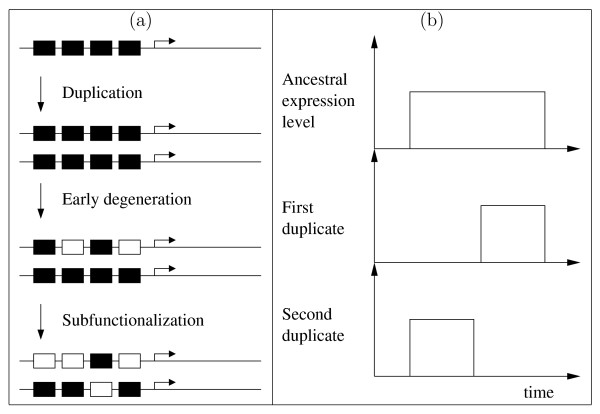
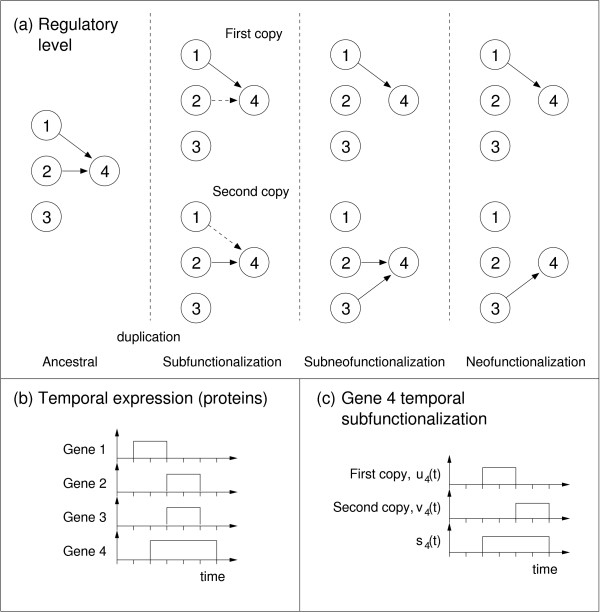
Background: The duplication-degeneration-complementation (DDC) model has been proposed as an explanation for the unexpectedly high retention of duplicate genes. The hypothesis proposes that, following gene duplication, the two gene copies degenerate to perform complementary functions that jointly match that of the single ancestral gene, a process also known as subfunctionalization. We distinguish between subfunctionalization at the regulatory level and at the product level (e.g within temporal or spatial expression domains).
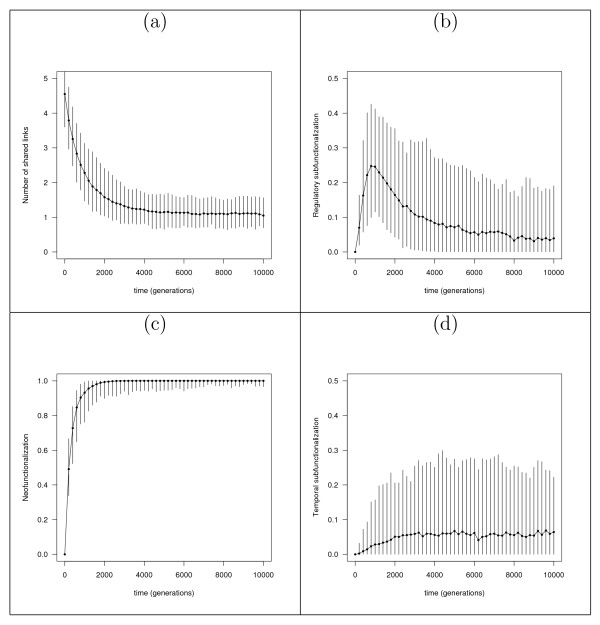
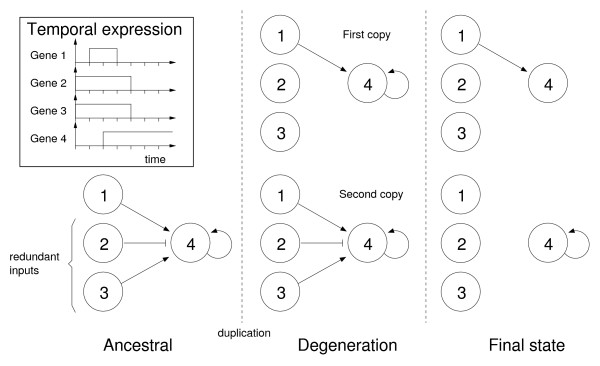
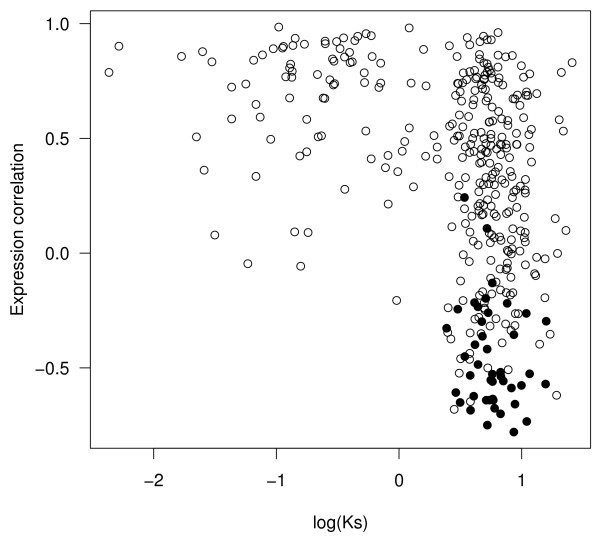
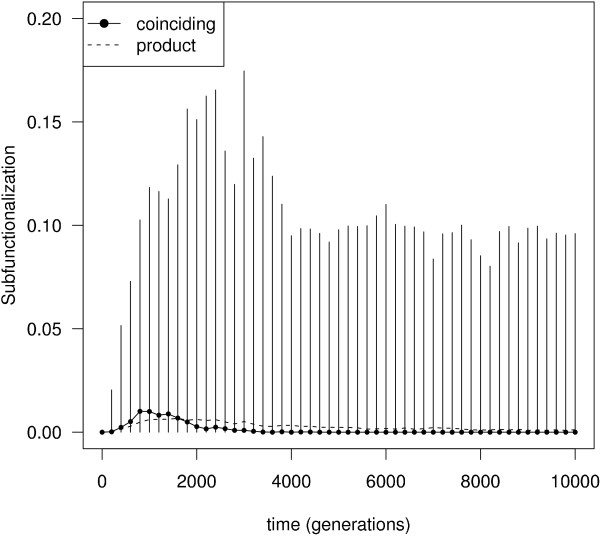
Results: In contrast to what is expected under the DDC model, we use in silico modeling to show that regulatory subfunctionalization is expected to peak and then decrease significantly. At the same time, neofunctionalization (recruitment of novel interactions) increases monotonically, eventually affecting the regulatory elements of the majority of genes. Furthermore, since this process occurs under conditions of stabilizing selection, there is no need to invoke positive selection. At the product level, the frequency of subfunctionalization is no higher than would be expected by chance, a finding that was corroborated using yeast microarray time-course data. We also find that product subfunctionalization is not necessarily caused by regulatory subfunctionalization.
Conclusion: Our results suggest a more complex picture of post-duplication evolution in which subfunctionalization plays only a partial role in conjunction with redundancy and neofunctionalization. We argue that this behavior is a consequence of the high evolutionary plasticity in gene networks.
Figures
References
-
- Hughes MK, Hughes AL. Evolution of duplicate genes in a tetraploid animal, Xenopus laevis. Mol Biol Evol. 1993;10:1360–1369. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
