

Genomic regulatory blocks underlie extensive microsynteny conservation in insects
- PMID: 17989259
- PMCID: PMC2099597
- DOI: 10.1101/gr.6669607
Genomic regulatory blocks underlie extensive microsynteny conservation in insects
Abstract
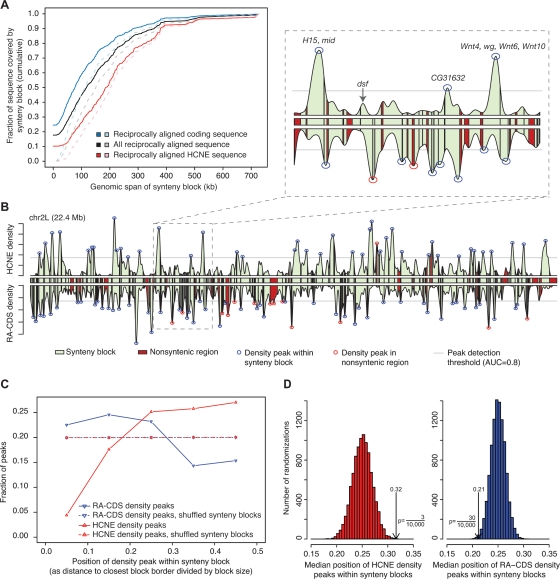
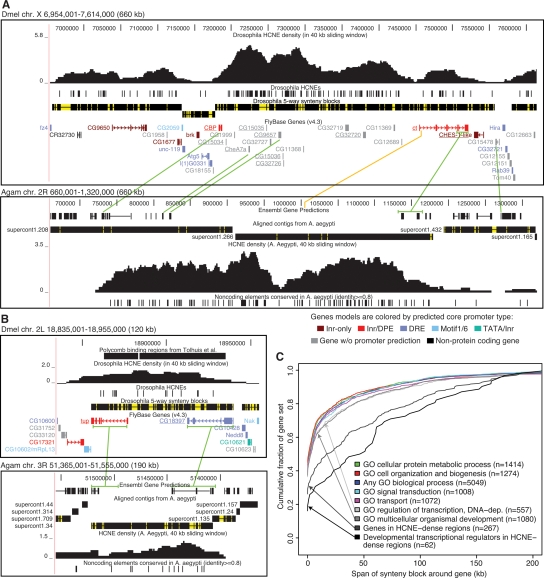
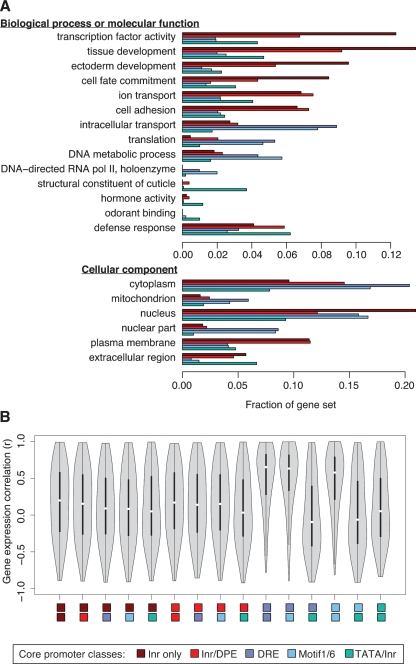
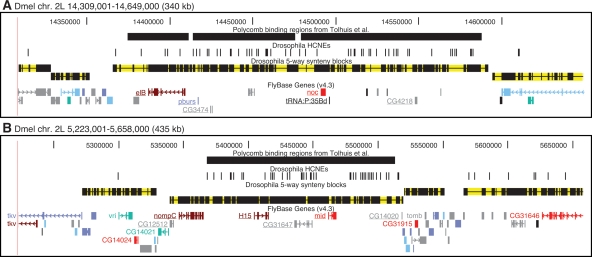
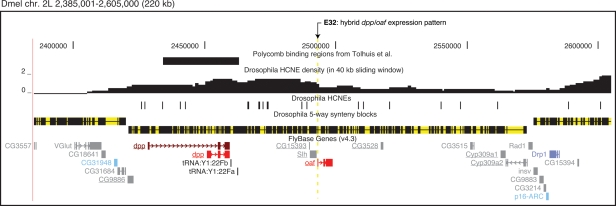
Insect genomes contain larger blocks of conserved gene order (microsynteny) than would be expected under a random breakage model of chromosome evolution. We present evidence that microsynteny has been retained to keep large arrays of highly conserved noncoding elements (HCNEs) intact. These arrays span key developmental regulatory genes, forming genomic regulatory blocks (GRBs). We recently described GRBs in vertebrates, where most HCNEs function as enhancers and HCNE arrays specify complex expression programs of their target genes. Here we present a comparison of five Drosophila genomes showing that HCNE density peaks centrally in large synteny blocks containing multiple genes. Besides developmental regulators that are likely targets of HCNE enhancers, HCNE arrays often span unrelated neighboring genes. We describe differences in core promoters between the target genes and the unrelated genes that offer an explanation for the differences in their responsiveness to enhancers. We show examples of a striking correspondence between boundaries of synteny blocks, HCNE arrays, and Polycomb binding regions, confirming that the synteny blocks correspond to regulatory domains. Although few noncoding elements are highly conserved between Drosophila and the malaria mosquito Anopheles gambiae, we find that A. gambiae regions orthologous to Drosophila GRBs contain an equivalent distribution of noncoding elements highly conserved in the yellow fever mosquito Aëdes aegypti and coincide with regions of ancient microsynteny between Drosophila and mosquitoes. The structural and functional equivalence between insect and vertebrate GRBs marks them as an ancient feature of metazoan genomes and as a key to future studies of development and gene regulation.
Figures
References
-
- Bailey P.J., Klos J.M., Andersson E., Karlen M., Kallstrom M., Ponjavic J., Muhr J., Lenhard B., Sandelin A., Ericson J., Klos J.M., Andersson E., Karlen M., Kallstrom M., Ponjavic J., Muhr J., Lenhard B., Sandelin A., Ericson J., Andersson E., Karlen M., Kallstrom M., Ponjavic J., Muhr J., Lenhard B., Sandelin A., Ericson J., Karlen M., Kallstrom M., Ponjavic J., Muhr J., Lenhard B., Sandelin A., Ericson J., Kallstrom M., Ponjavic J., Muhr J., Lenhard B., Sandelin A., Ericson J., Ponjavic J., Muhr J., Lenhard B., Sandelin A., Ericson J., Muhr J., Lenhard B., Sandelin A., Ericson J., Lenhard B., Sandelin A., Ericson J., Sandelin A., Ericson J., Ericson J. A global genomic transcriptional code associated with CNS-expressed genes. Exp. Cell Res. 2006;312:3108–3119. - PubMed
-
- Bejerano G., Pheasant M., Makunin I., Stephen S., Kent W.J., Mattick J.S., Haussler D., Pheasant M., Makunin I., Stephen S., Kent W.J., Mattick J.S., Haussler D., Makunin I., Stephen S., Kent W.J., Mattick J.S., Haussler D., Stephen S., Kent W.J., Mattick J.S., Haussler D., Kent W.J., Mattick J.S., Haussler D., Mattick J.S., Haussler D., Haussler D. Ultraconserved elements in the human genome. Science. 2004;304:1321–1325. - PubMed
-
- Boyer L.A., Plath K., Zeitlinger J., Brambrink T., Medeiros L.A., Lee T.I., Levine S.S., Wernig M., Tajonar A., Ray M.K., Plath K., Zeitlinger J., Brambrink T., Medeiros L.A., Lee T.I., Levine S.S., Wernig M., Tajonar A., Ray M.K., Zeitlinger J., Brambrink T., Medeiros L.A., Lee T.I., Levine S.S., Wernig M., Tajonar A., Ray M.K., Brambrink T., Medeiros L.A., Lee T.I., Levine S.S., Wernig M., Tajonar A., Ray M.K., Medeiros L.A., Lee T.I., Levine S.S., Wernig M., Tajonar A., Ray M.K., Lee T.I., Levine S.S., Wernig M., Tajonar A., Ray M.K., Levine S.S., Wernig M., Tajonar A., Ray M.K., Wernig M., Tajonar A., Ray M.K., Tajonar A., Ray M.K., Ray M.K., et al. Polycomb complexes repress developmental regulators in murine embryonic stem cells. Nature. 2006;441:349–353. - PubMed
-
- Brudno M., Malde S., Poliakov A., Do C.B., Couronne O., Dubchak I., Batzoglou S., Malde S., Poliakov A., Do C.B., Couronne O., Dubchak I., Batzoglou S., Poliakov A., Do C.B., Couronne O., Dubchak I., Batzoglou S., Do C.B., Couronne O., Dubchak I., Batzoglou S., Couronne O., Dubchak I., Batzoglou S., Dubchak I., Batzoglou S., Batzoglou S. Glocal alignment: Finding rearrangements during alignment. Bioinformatics. 2003;19:i54–i62. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases