

Neural ensembles in CA3 transiently encode paths forward of the animal at a decision point
- PMID: 17989284
- PMCID: PMC6673267
- DOI: 10.1523/JNEUROSCI.3761-07.2007
Neural ensembles in CA3 transiently encode paths forward of the animal at a decision point
Abstract
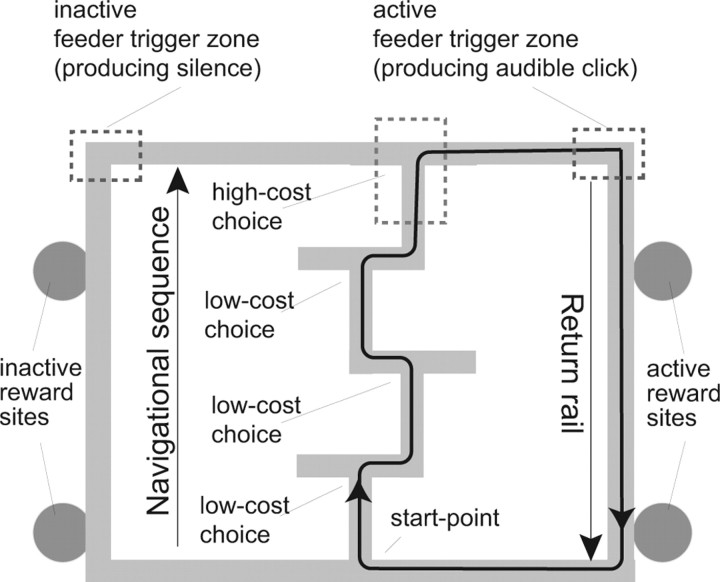
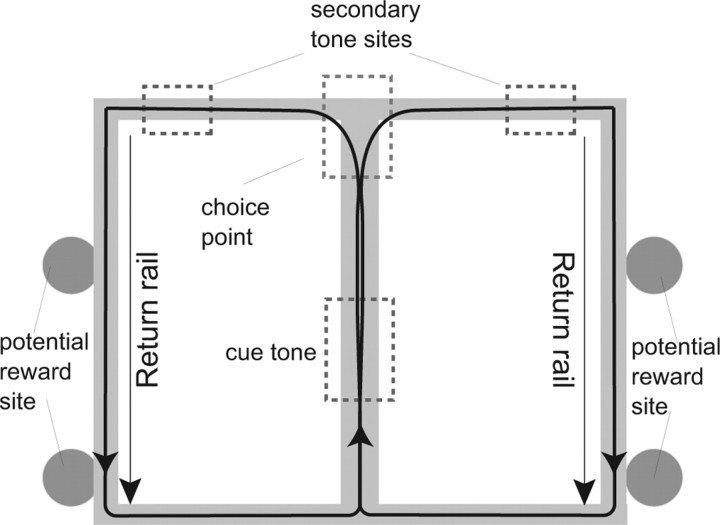
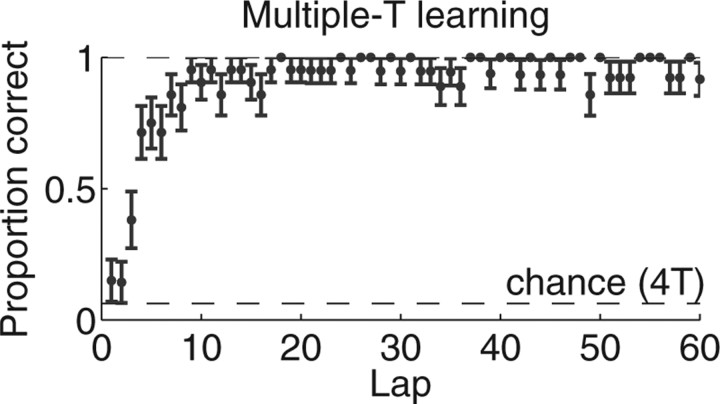
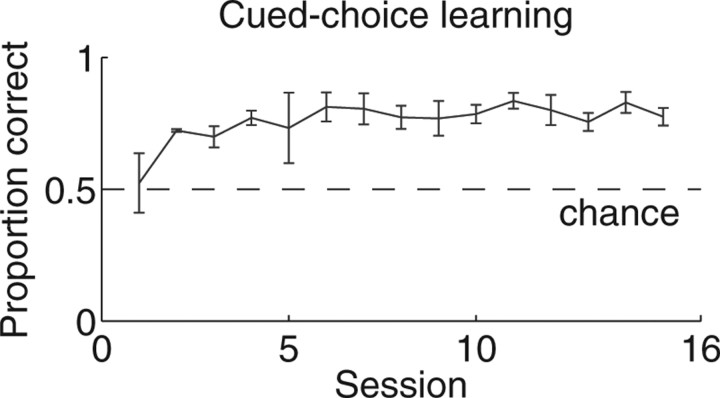
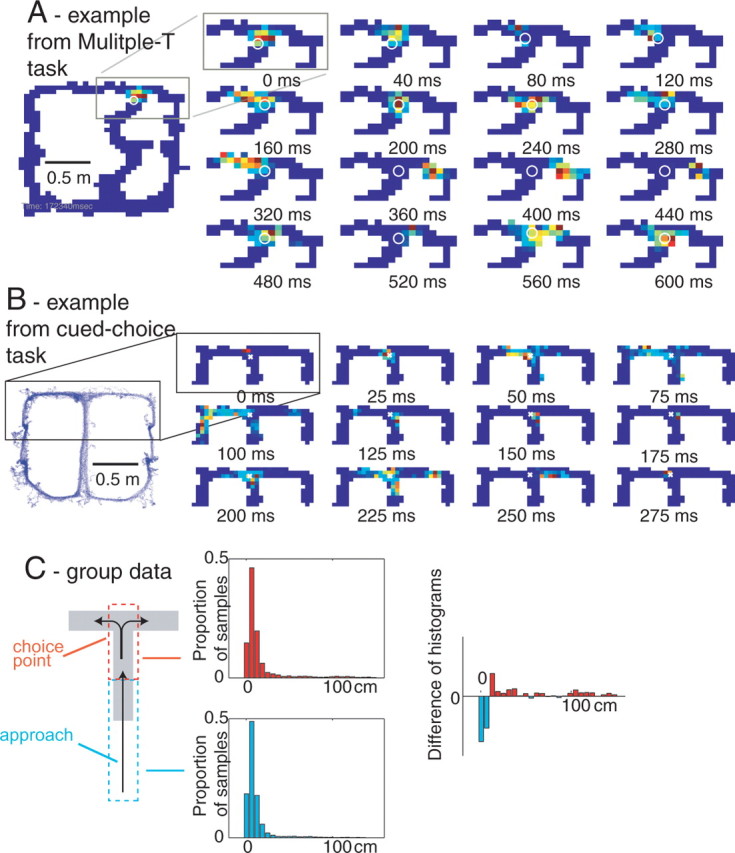
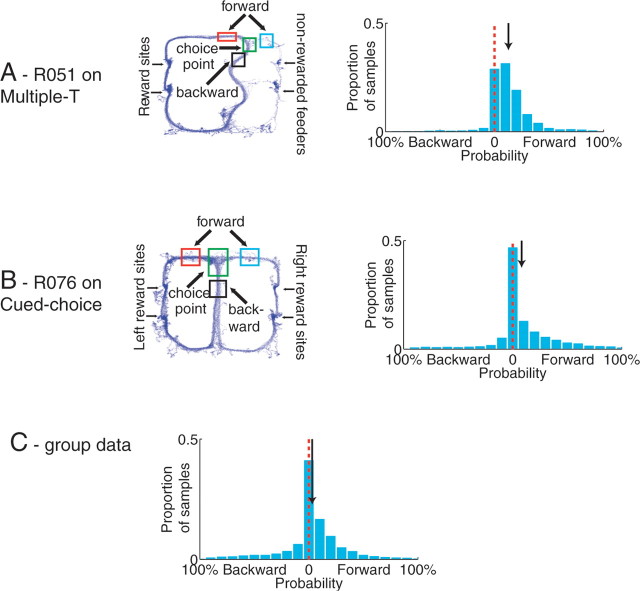
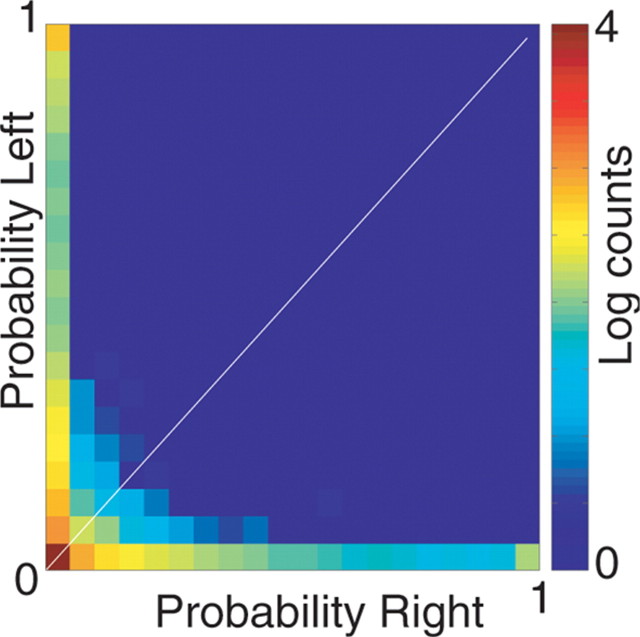
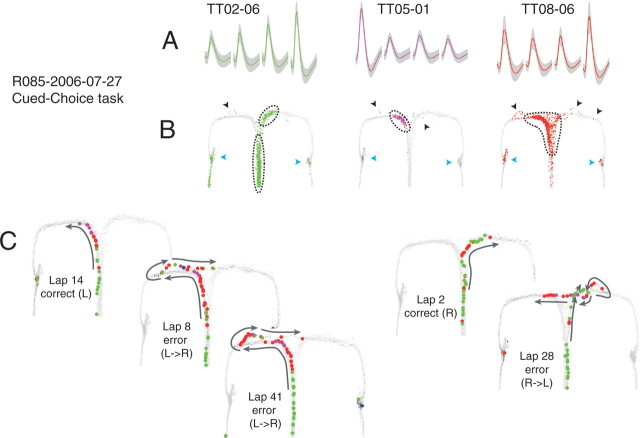
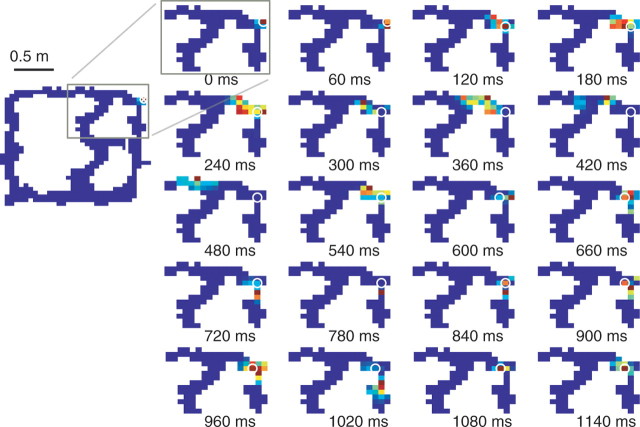
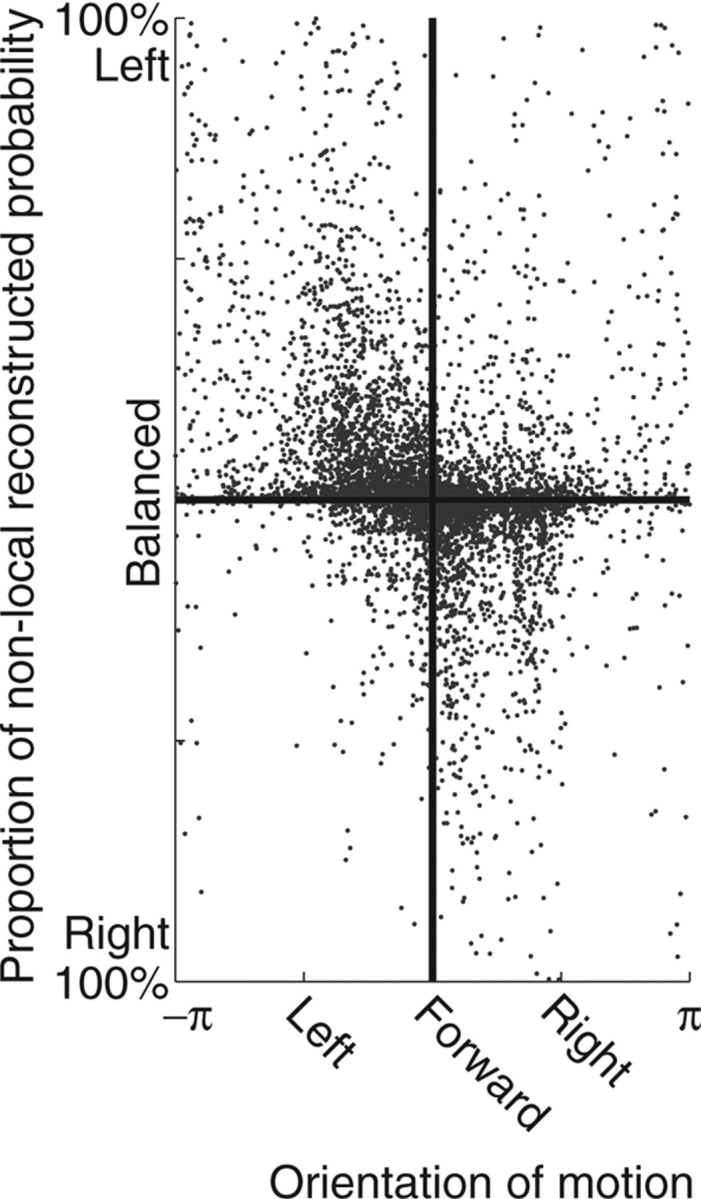
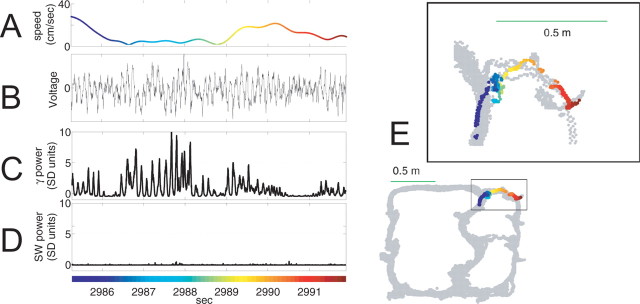
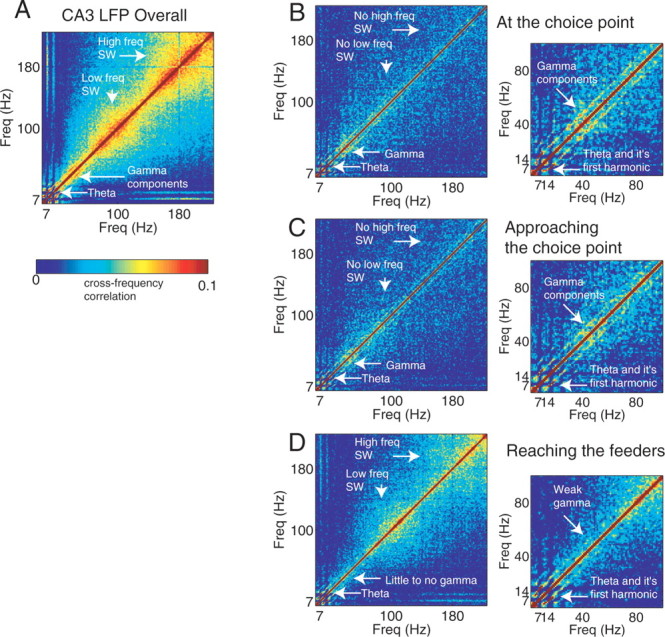
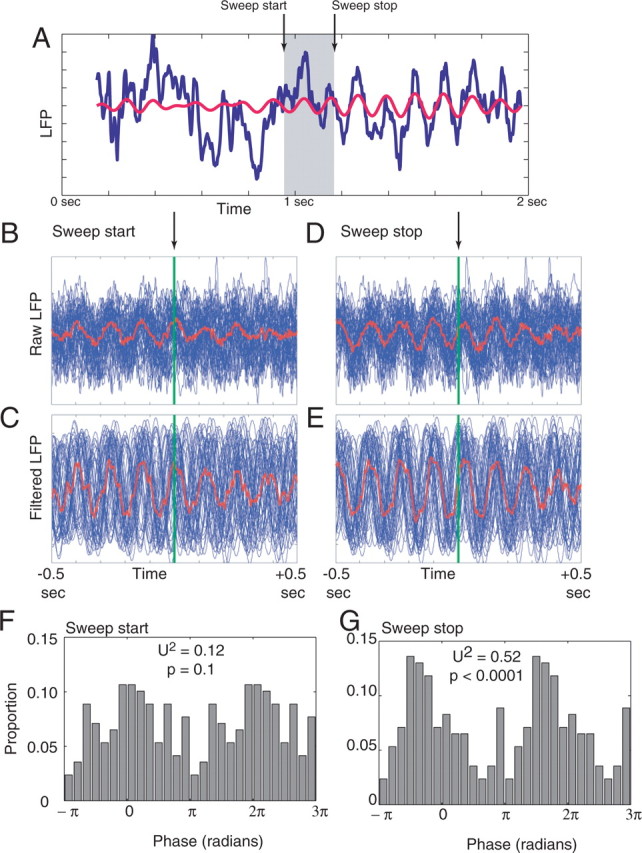
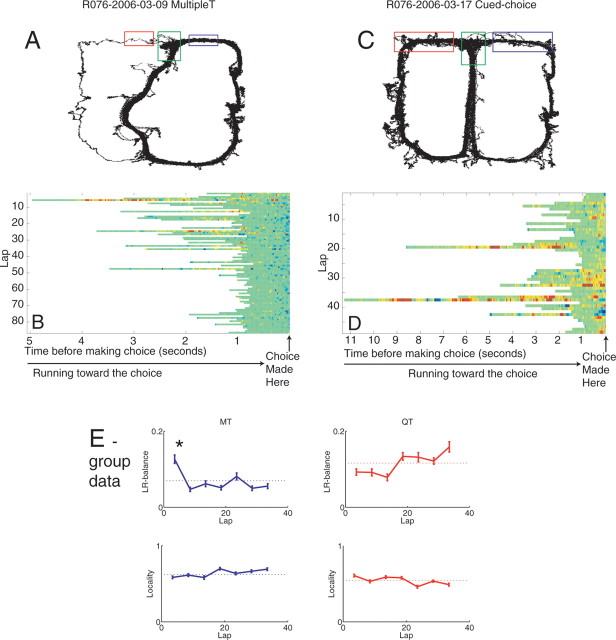
Neural ensembles were recorded from the CA3 region of rats running on T-based decision tasks. Examination of neural representations of space at fast time scales revealed a transient but repeatable phenomenon as rats made a decision: the location reconstructed from the neural ensemble swept forward, first down one path and then the other. Estimated representations were coherent and preferentially swept ahead of the animal rather than behind the animal, implying it represented future possibilities rather than recently traveled paths. Similar phenomena occurred at other important decisions (such as in recovery from an error). Local field potentials from these sites contained pronounced theta and gamma frequencies, but no sharp wave frequencies. Forward-shifted spatial representations were influenced by task demands and experience. These data suggest that the hippocampus does not represent space as a passive computation, but rather that hippocampal spatial processing is an active process likely regulated by cognitive mechanisms.
Figures

References
-
- Ainge JA, van der Meer MA, Wood ER. Disparity between sequence-dependent hippocampal activity and hippocampal lesion effects on a continuous t-maze task. Soc Neurosci Abstr. 2005;31:776–13.
-
- Ainge JA, van der Meer MA, Langston RF, Wood ER. Exploring the role of context-dependent hippocampal activity in spatial alternation behavior. Hippocampus. 2007b;17:988–1002. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous