

A bifunctional locus (BIO3-BIO1) required for biotin biosynthesis in Arabidopsis
- PMID: 17993549
- PMCID: PMC2230573
- DOI: 10.1104/pp.107.107409
A bifunctional locus (BIO3-BIO1) required for biotin biosynthesis in Arabidopsis
Abstract
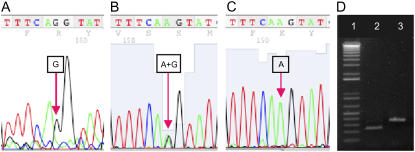
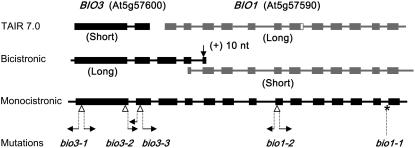
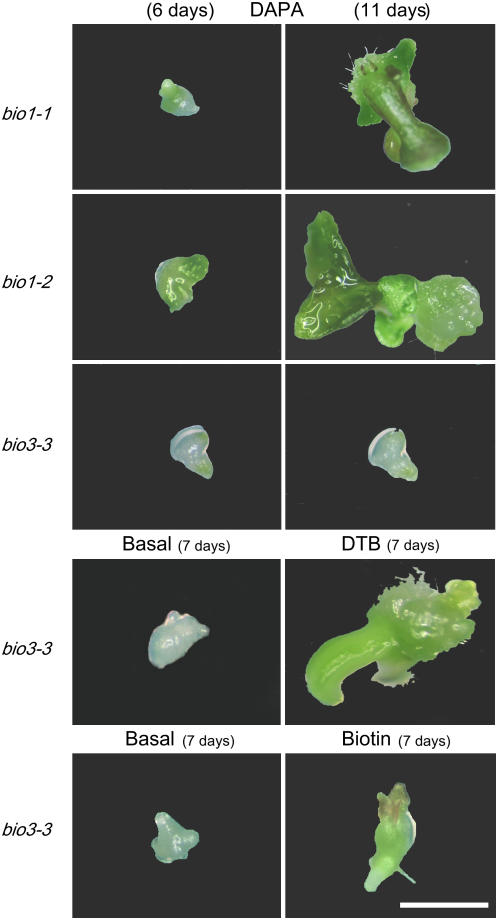
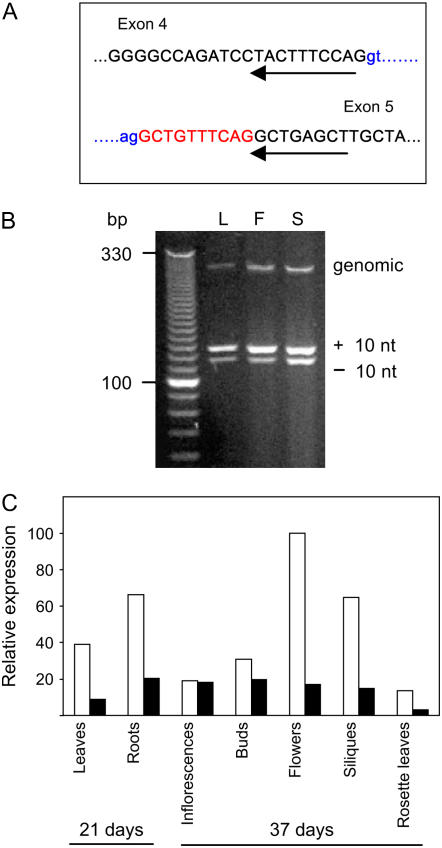
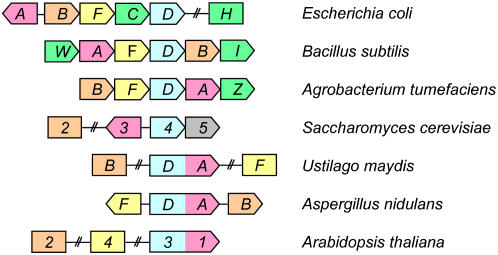
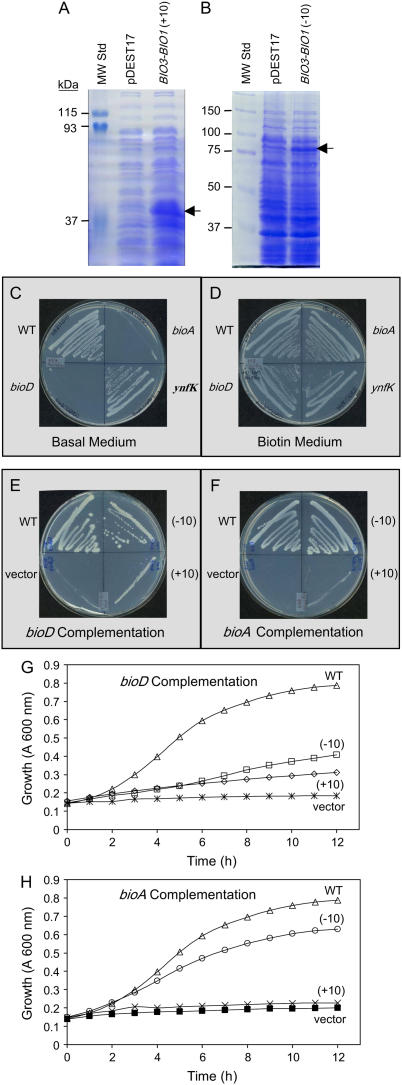
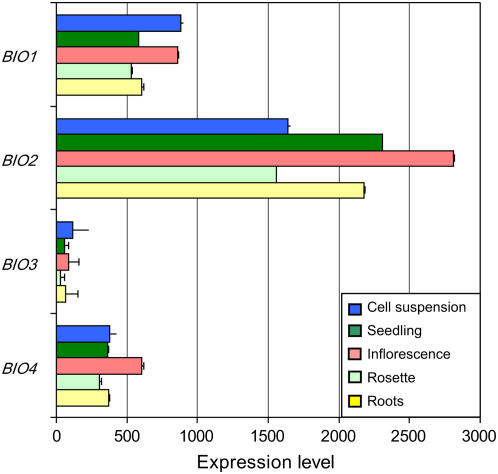
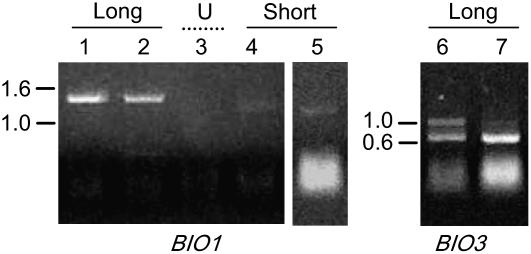
We identify here the Arabidopsis (Arabidopsis thaliana) gene encoding the third enzyme in the biotin biosynthetic pathway, dethiobiotin synthetase (BIO3; At5g57600). This gene is positioned immediately upstream of BIO1, which is known to be associated with the second reaction in the pathway. Reverse genetic analysis demonstrates that bio3 insertion mutants have a similar phenotype to the bio1 and bio2 auxotrophs identified using forward genetic screens for arrested embryos rescued on enriched nutrient medium. Unexpectedly, bio3 and bio1 mutants define a single genetic complementation group. Reverse transcription-polymerase chain reaction analysis demonstrates that separate BIO3 and BIO1 transcripts and two different types of chimeric BIO3-BIO1 transcripts are produced. Consistent with genetic data, one of the fused transcripts is monocistronic and encodes a bifunctional fusion protein. A splice variant is bicistronic, with distinct but overlapping reading frames. The dual functionality of the monocistronic transcript was confirmed by complementing the orthologous auxotrophs of Escherichia coli (bioD and bioA). BIO3-BIO1 transcripts from other plants provide further evidence for differential splicing, existence of a fusion protein, and localization of both enzymatic reactions to mitochondria. In contrast to most biosynthetic enzymes in eukaryotes, which are encoded by genes dispersed throughout the genome, biotin biosynthesis in Arabidopsis provides an intriguing example of a bifunctional locus that catalyzes two sequential reactions in the same metabolic pathway. This complex locus exhibits several unusual features that distinguish it from biotin operons in bacteria and from other genes known to encode bifunctional enzymes in plants.
Figures

Similar articles
-
Biochemical and structural characterization of the Arabidopsis bifunctional enzyme dethiobiotin synthetase-diaminopelargonic acid aminotransferase: evidence for substrate channeling in biotin synthesis.Plant Cell. 2012 Apr;24(4):1608-25. doi: 10.1105/tpc.112.097675. Epub 2012 Apr 30. Plant Cell. 2012. PMID: 22547782 Free PMC article.
-
Cadaverine regulates biotin synthesis to modulate primary root growth in Arabidopsis.Plant J. 2021 Sep;107(5):1283-1298. doi: 10.1111/tpj.15417. Epub 2021 Aug 12. Plant J. 2021. PMID: 34250670 Free PMC article.
-
Complementation of an Arabidopsis thaliana biotin auxotroph with an Escherichia coli biotin biosynthetic gene.Mol Gen Genet. 1996 Jun 12;251(3):261-6. doi: 10.1007/BF02172516. Mol Gen Genet. 1996. PMID: 8676868
-
Genomic organization of the biotin biosynthetic genes of coryneform bacteria: cloning and sequencing of the bioA-bioD genes from Brevibacterium flavum.DNA Seq. 1993;4(3):177-84. doi: 10.3109/10425179309015630. DNA Seq. 1993. PMID: 8161820
-
Characterization of the biotin biosynthesis pathway in Saccharomyces cerevisiae and evidence for a cluster containing BIO5, a novel gene involved in vitamer uptake.Gene. 1999 May 17;232(1):43-51. doi: 10.1016/s0378-1119(99)00117-1. Gene. 1999. PMID: 10333520
Cited by
-
SUCROSE TRANSPORTER 5 supplies Arabidopsis embryos with biotin and affects triacylglycerol accumulation.Plant J. 2013 Feb;73(3):392-404. doi: 10.1111/tpj.12037. Epub 2012 Dec 31. Plant J. 2013. PMID: 23031218 Free PMC article.
-
Natural variation in sensitivity to a loss of chloroplast translation in Arabidopsis.Plant Physiol. 2014 Dec;166(4):2013-27. doi: 10.1104/pp.114.249052. Epub 2014 Oct 21. Plant Physiol. 2014. PMID: 25336520 Free PMC article.
-
Operons.Cell Mol Life Sci. 2009 Dec;66(23):3755-75. doi: 10.1007/s00018-009-0114-3. Epub 2009 Aug 7. Cell Mol Life Sci. 2009. PMID: 19662496 Free PMC article. Review.
-
A newly discovered function of peroxisomes: involvement in biotin biosynthesis.Plant Signal Behav. 2012 Dec;7(12):1589-93. doi: 10.4161/psb.22405. Epub 2012 Oct 16. Plant Signal Behav. 2012. PMID: 23073000 Free PMC article. Review.
-
Cross-exchange of B-vitamins underpins a mutualistic interaction between Ostreococcus tauri and Dinoroseobacter shibae.ISME J. 2019 Feb;13(2):334-345. doi: 10.1038/s41396-018-0274-y. Epub 2018 Sep 18. ISME J. 2019. PMID: 30228381 Free PMC article.
References
-
- Alonso JM, Stepanova AN, Leisse TJ, Kim CJ, Chen H, Shinn P, Stevenson DK, Zimmerman J, Barajas P, Cheuk R, et al (2003) Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301 653–657 - PubMed
-
- Arnal N, Alban C, Quadrado M, Grandjean O, Mireau H (2006) The Arabidopsis Bio2 protein requires mitochondrial targeting for activity. Plant Mol Biol 62 471–479 - PubMed
-
- Ashkenazi T, Pinkert D, Nudelman A, Widberg A, Wexler B, Wittenbach V, Flint D, Nudelman A (2007) Aryl chain analogues of the biotin vitamers as potential herbicides. Part 3. Pest Manag Sci 63 974–1001 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
