

Adherence to macrophages in erythroblastic islands enhances erythroblast proliferation and increases erythrocyte production by a different mechanism than erythropoietin
- PMID: 17993612
- PMCID: PMC2214751
- DOI: 10.1182/blood-2007-06-098178
Adherence to macrophages in erythroblastic islands enhances erythroblast proliferation and increases erythrocyte production by a different mechanism than erythropoietin
Abstract
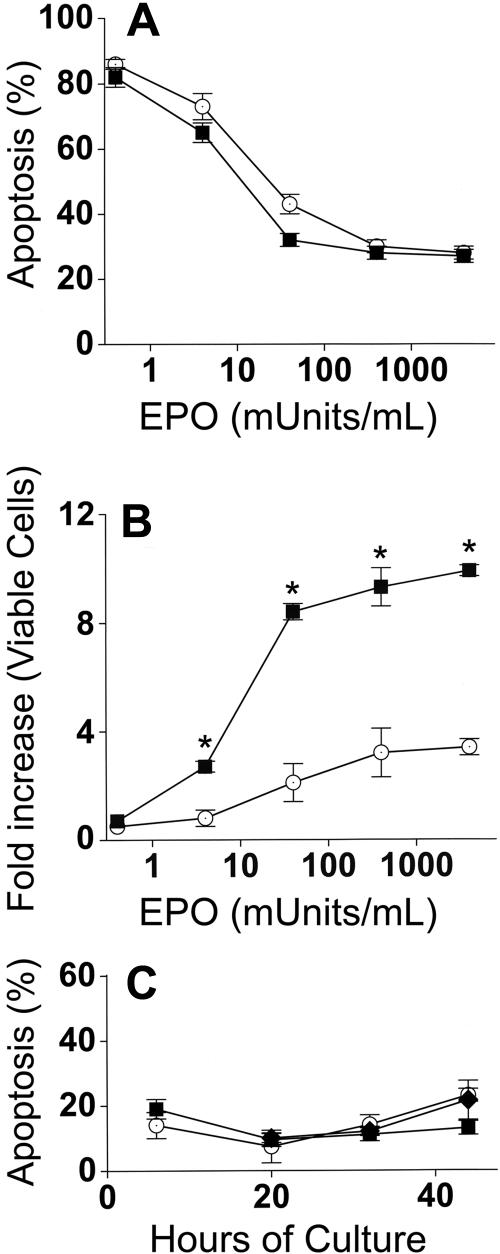
Erythroblasts adhere to central macrophages forming erythroblastic islands in hematopoietic tissues, but the function of these islands is not understood. Murine erythroblastic islands were reconstituted in vitro with macrophages and developmentally synchronous proerythroblasts. Erythroblasts cocultured with macrophages proliferated 3-fold greater than erythroblasts cultured alone. Direct contact with the macrophages was necessary for this enhanced erythroblast proliferation, which resulted from decreased transit time in the G(0)/G(1) phase of cell cycle. Increased erythroblast proliferation in erythroblastic islands occurred over a wide range of erythropoietin concentrations and was the result of a mechanism different from the antiapoptotic effect of erythropoietin. Erythroblasts adherent to macrophages had slightly delayed enucleation, but otherwise differentiation was similar to erythroblasts cultured alone or those that became nonadherent in cocultures. These results suggest a mechanism for the development of anemias associated with abnormal macrophage function and for reduced responsiveness of those anemias to erythropoietin therapy.
Figures


 ), nonadherent erythroblasts in cocultures (■), and in control cultures (○). Results are the means plus or minus SE from 24 separate experiments; increased accumulations of erythroblasts in cocultures compared with control cultures were significant (*P < .001) at all times after 6 hours.
), nonadherent erythroblasts in cocultures (■), and in control cultures (○). Results are the means plus or minus SE from 24 separate experiments; increased accumulations of erythroblasts in cocultures compared with control cultures were significant (*P < .001) at all times after 6 hours.



References
-
- Chasis JA. Erythroblastic islands: specialized microenvironmental niches for erythropoiesis. Curr Opin Hematol. 2006;13:137–141. - PubMed
-
- Le Charpentier Y, Prenant M. [Isolation of erythroblastic islands. Study by optical and scanning electron microscopy (author's transl)]. Nouv Rev Fr Hematol. 1975;15:119–140. - PubMed
-
- Bessis M, Mize C, Prenant M. Erythropoiesis: comparison of in vivo and in vitro amplification. Blood Cells. 1978;4:155–174. - PubMed
-
- Hanspal M, Hanspal JS. The association of erythroblasts with macrophages promotes erythroid proliferation and maturation: a 30-kD heparin-binding protein is involved in this contact. Blood. 1994;84:3494–3504. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
