

Reactive oxygen species mediate Na+-induced SOS1 mRNA stability in Arabidopsis
- PMID: 17996020
- PMCID: PMC3128381
- DOI: 10.1111/j.1365-313X.2007.03364.x
Reactive oxygen species mediate Na+-induced SOS1 mRNA stability in Arabidopsis
Abstract
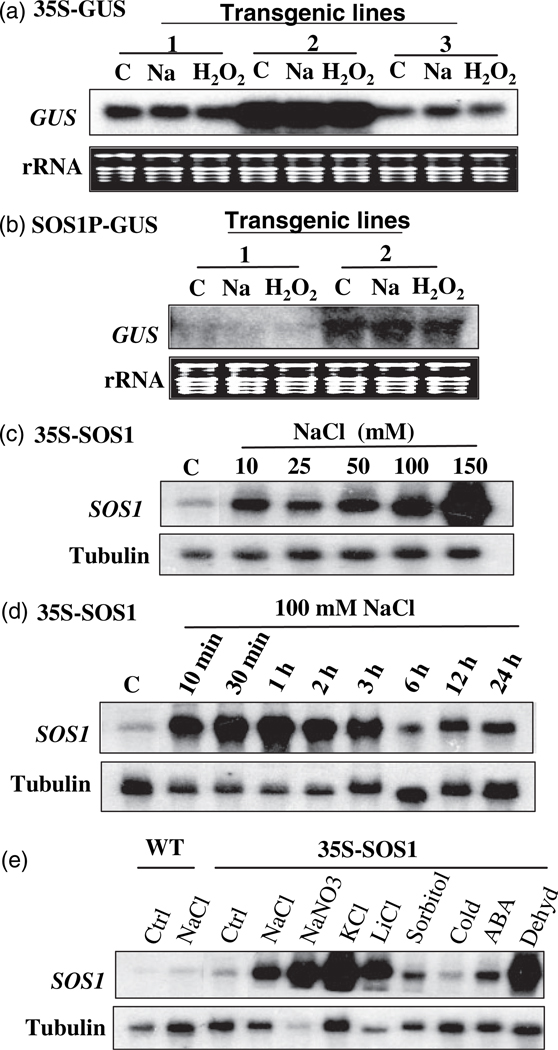
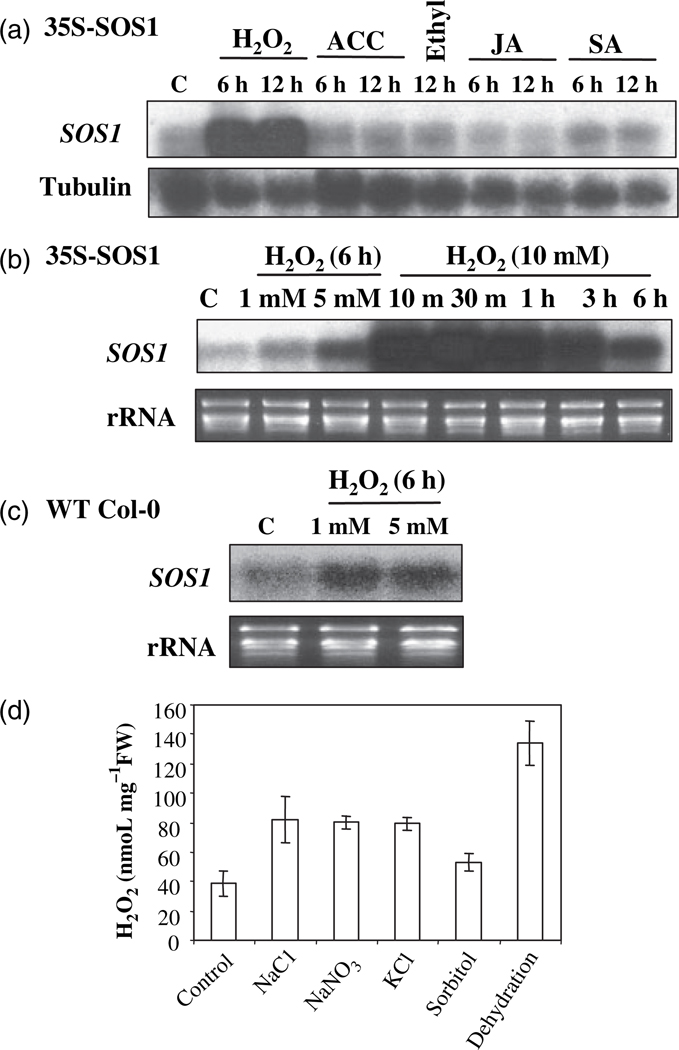
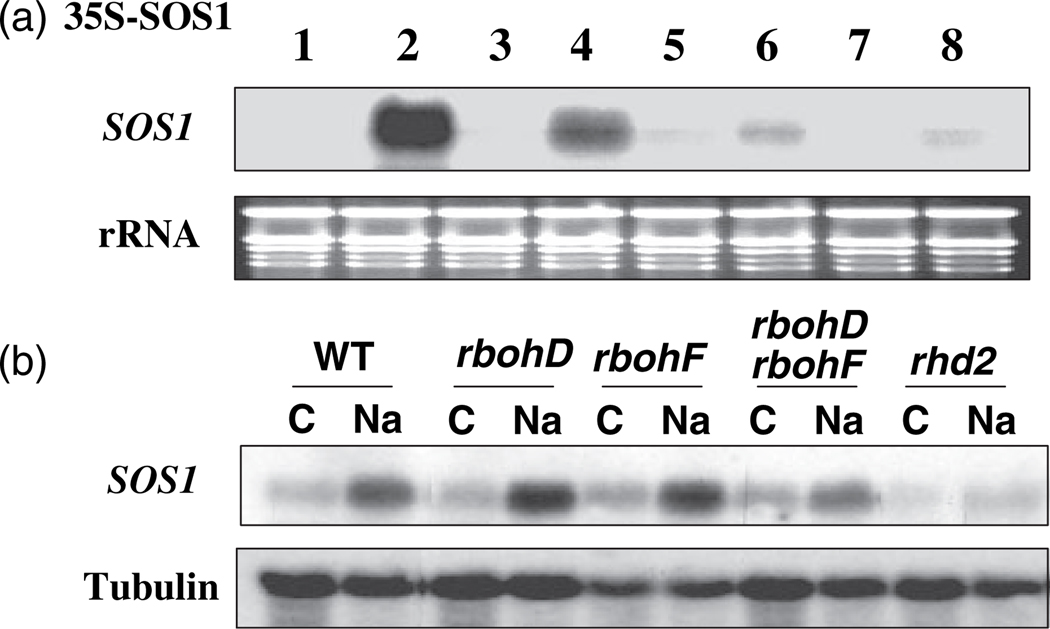
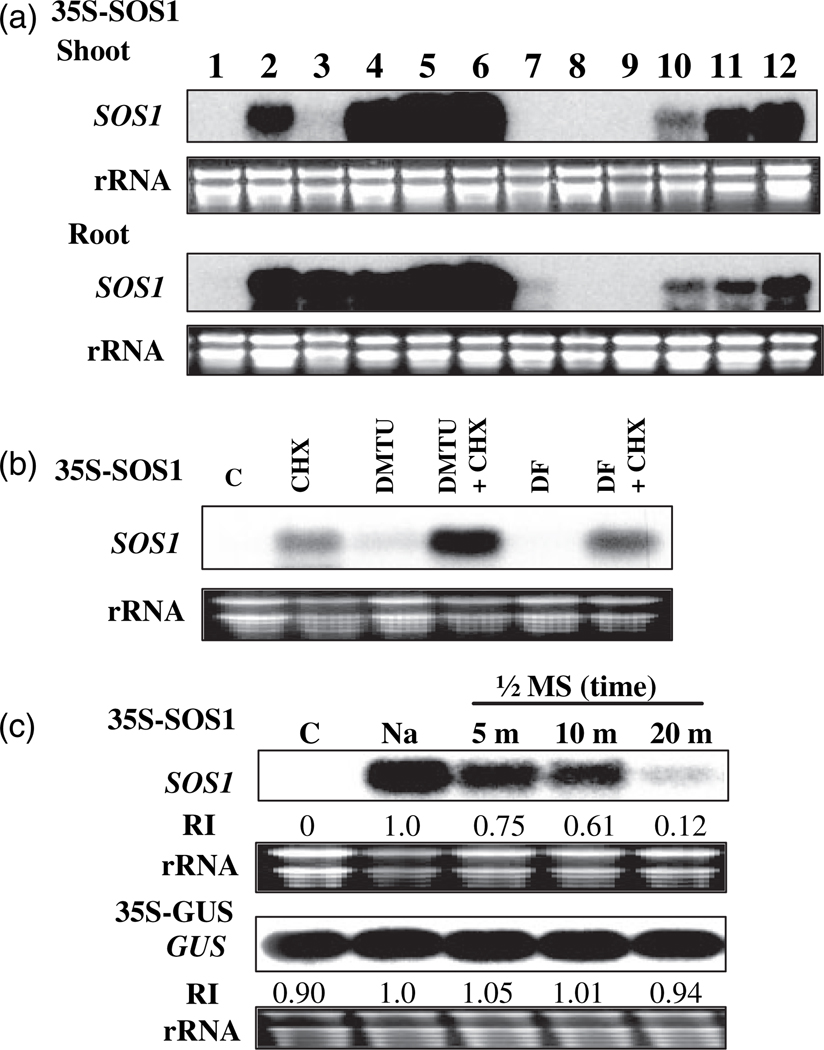
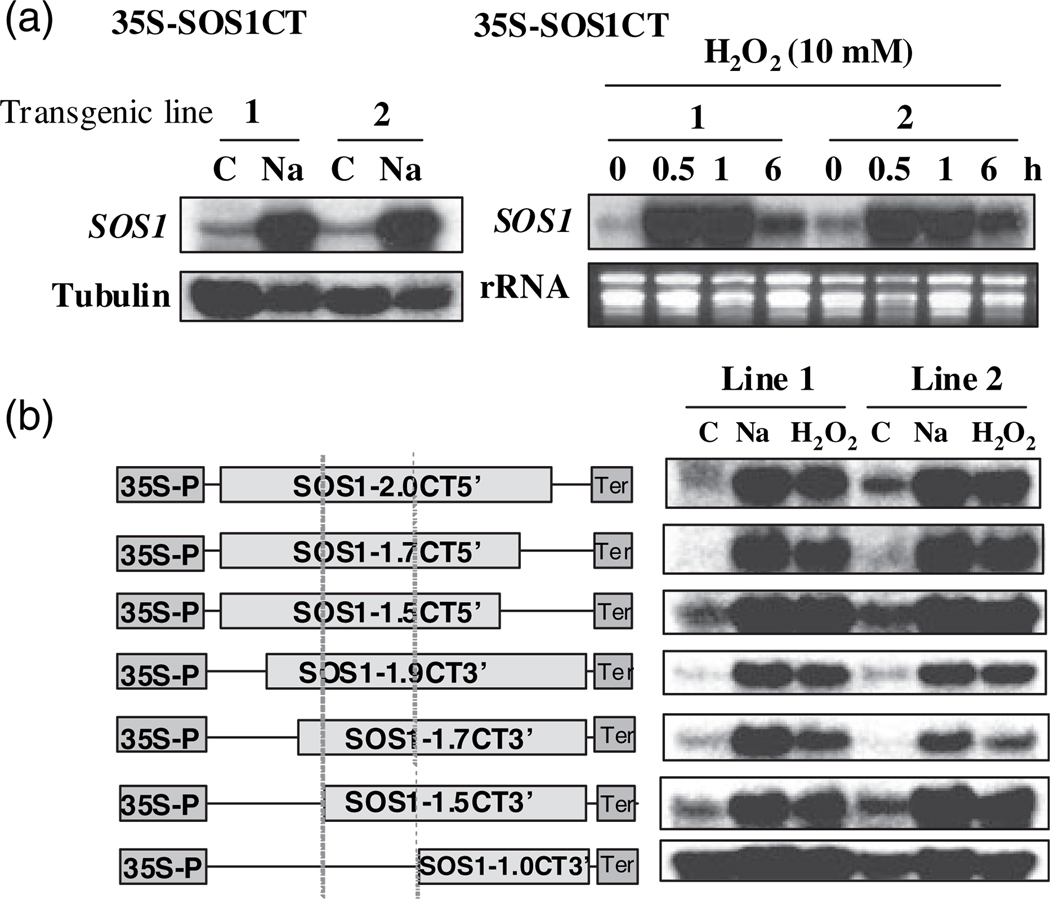
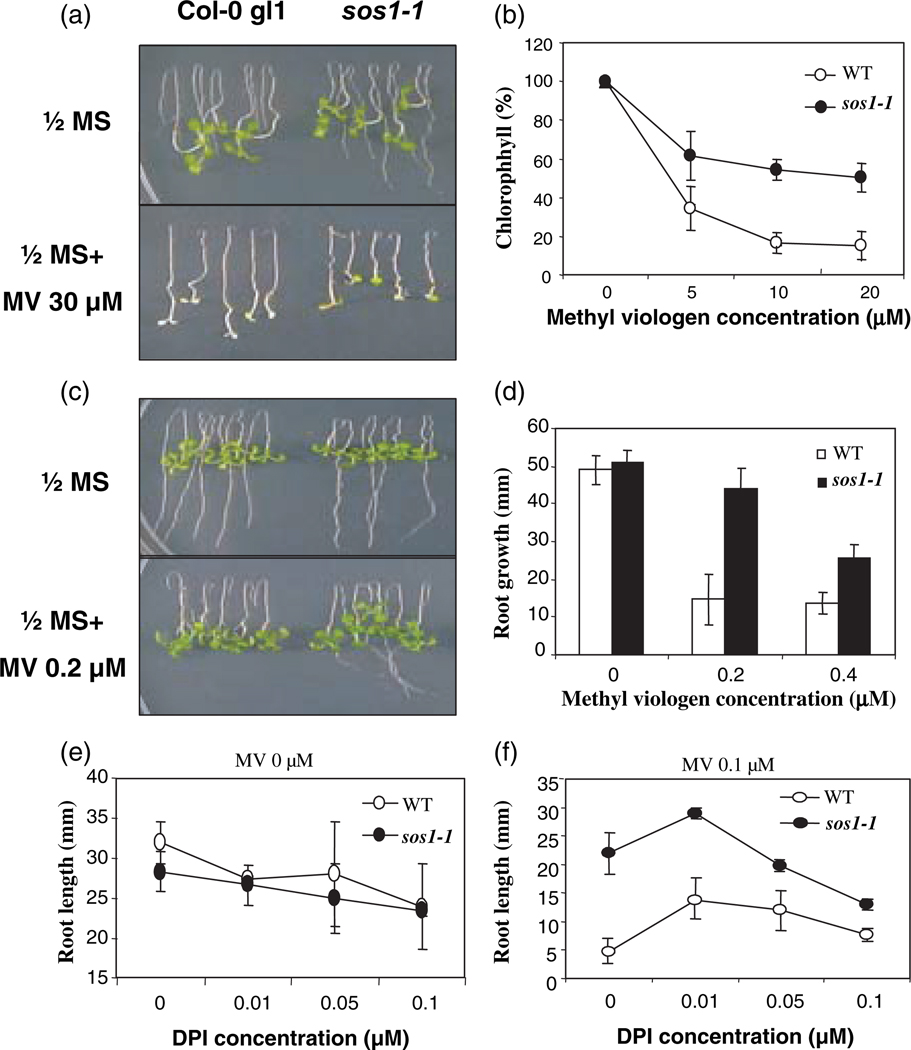
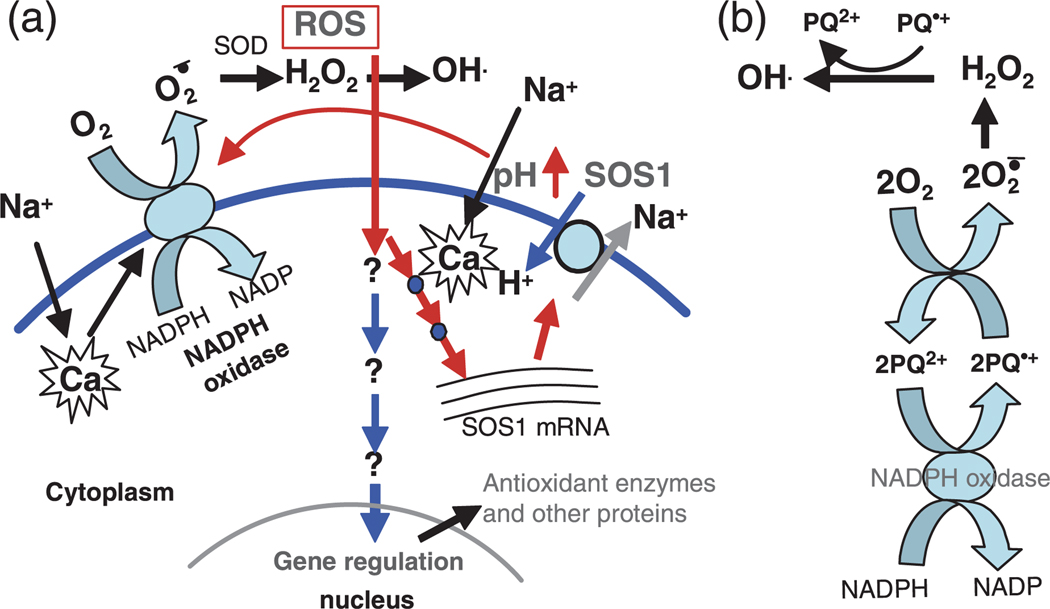
Salt Overly Sensitive 1 (SOS1), a plasma membrane Na+/H+ antiporter in Arabidopsis, is a salt tolerance determinant crucial for the maintenance of ion homeostasis in saline stress conditions. SOS1 mRNA is unstable at normal growth conditions, but its stability is substantially increased under salt stress and other ionic and dehydration stresses. In addition, H2O2 treatment increases the stability of SOS1 mRNA. SOS1 mRNA is inherently unstable and rapidly degraded with a half-life of approximately 10 min. Rapid decay of SOS1 mRNA requires new protein synthesis. Stress-induced SOS1 mRNA stability is mediated by reactive oxygen species (ROS). NADPH oxidase is also involved in the upregulation of SOS1 mRNA stability, presumably through the control of extracellular ROS production. The cis-element required for SOS1 mRNA instability resides in the 500-bp region within the 2.2 kb at the 3' end of the SOS1 mRNA. Furthermore, mutations in the SOS1 gene render sos1 mutants more tolerant to paraquat, a non-selective herbicide causing oxidative stress, indicating that SOS1 plays negative roles in tolerance of oxidative stress. A hypothetical model for the signaling pathway involving SOS1-mediated pH changes, NADPH oxidase activation, apoplastic ROS production and downstream signaling transduction is proposed, and the biological significance of ROS-mediated induction of SOS1 mRNA stability is discussed.
Figures
References
-
- Apel K, Hirt H. Reactive oxygen species: metabolism, oxidative stress, and signal transduction. Annu. Rev. Plant. Biol. 2004;55:373–399. - PubMed
-
- Bindschedler LV, Minibayeva F, Gardner SL, Gerrish C, Davis DR, Bolwell GP. Early signaling events in the apoplatic oxidative burst in suspension-cultured French bean cells involve cAMP and Ca2+ New Phytol. 2001;151:185–194. - PubMed
-
- Bolwell GP, Butt VS, Davies DR, Zimmerlin A. The origin of the oxidative burst in plants. Free Radic. Res. 1995;23:517–532. - PubMed
-
- Bolwell GP, Blee KA, Butt VS, Davies DR, Gardner SL, Gerrish C, Minibayeva F, Rowntree EG, Wojtaszek P. Recent advances in understanding the origin of the apoplastic oxidative burst in plant cells. Free Radic. Res. 1999;31 Suppl.:S137–S145. - PubMed
-
- Bolwell GP, Bindschedler LV, Blee KA, Butt VS, Davies DR, Gardner SL, Gerrish C, Minibayeva F. The apoplastic oxidative burst in response to biotic stress in plants: a three-component system. J. Exp. Bot. 2002;53:1367–1376. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
