

Molecular and functional mapping of EED motifs required for PRC2-dependent histone methylation
- PMID: 17997413
- PMCID: PMC2693252
- DOI: 10.1016/j.jmb.2007.10.040
Molecular and functional mapping of EED motifs required for PRC2-dependent histone methylation
Abstract
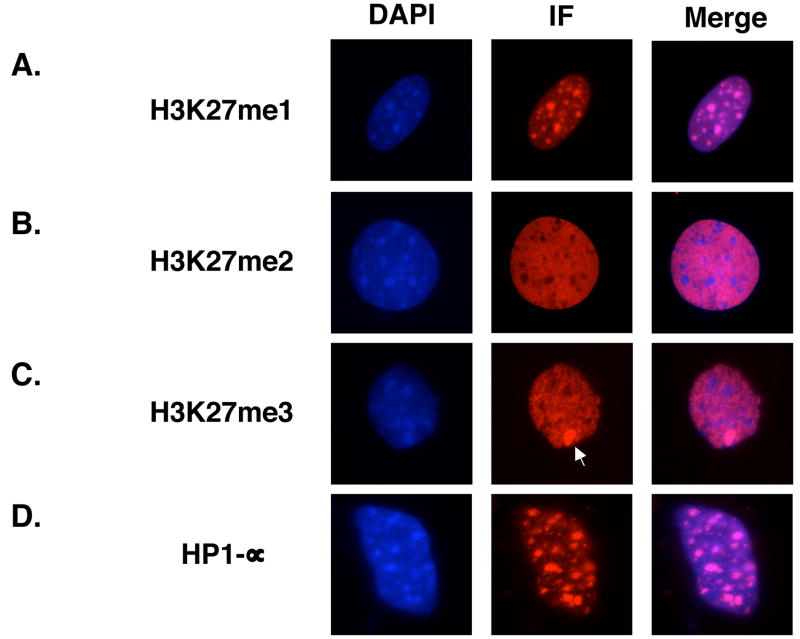
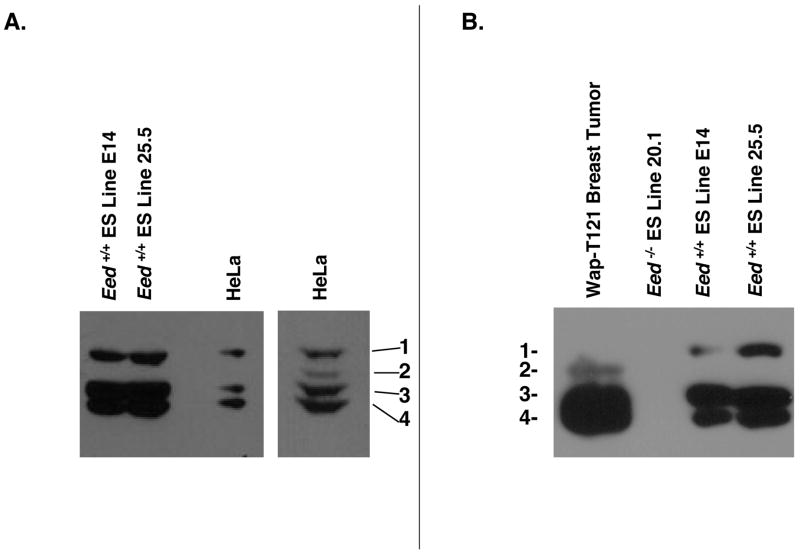
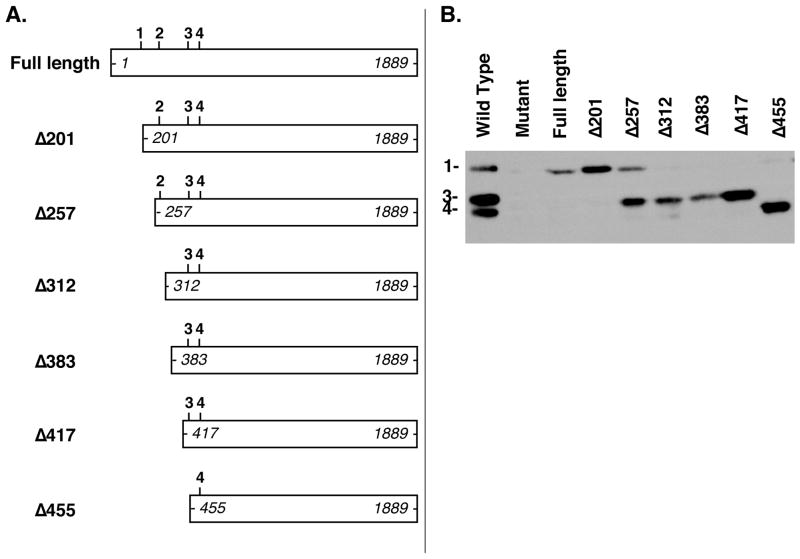
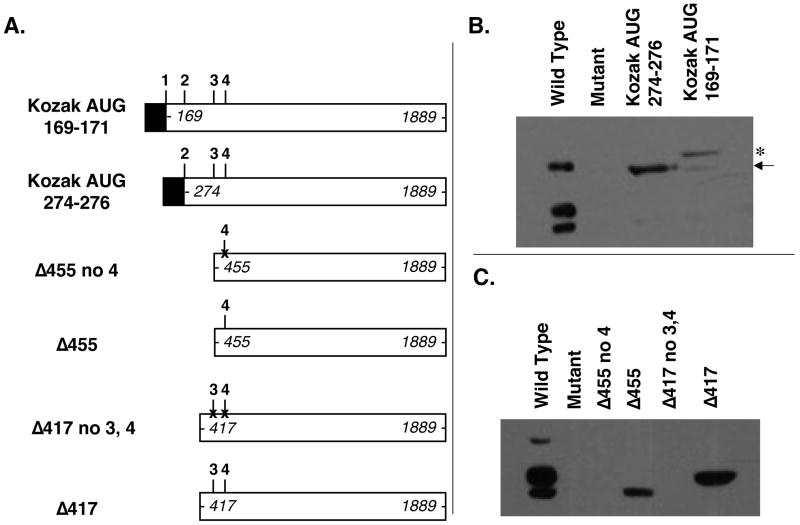
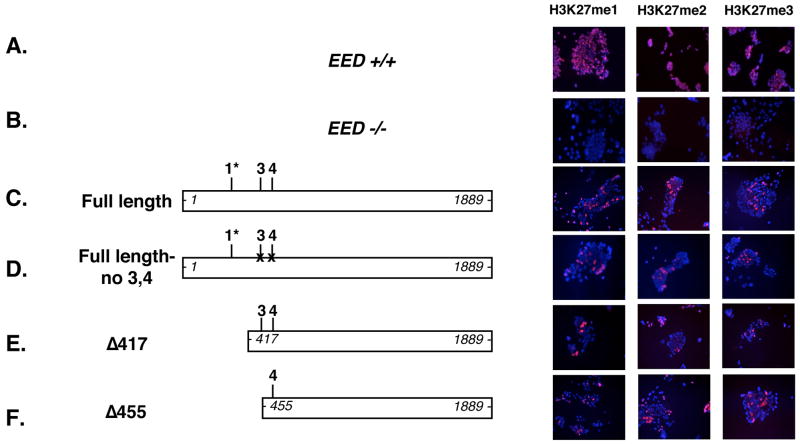
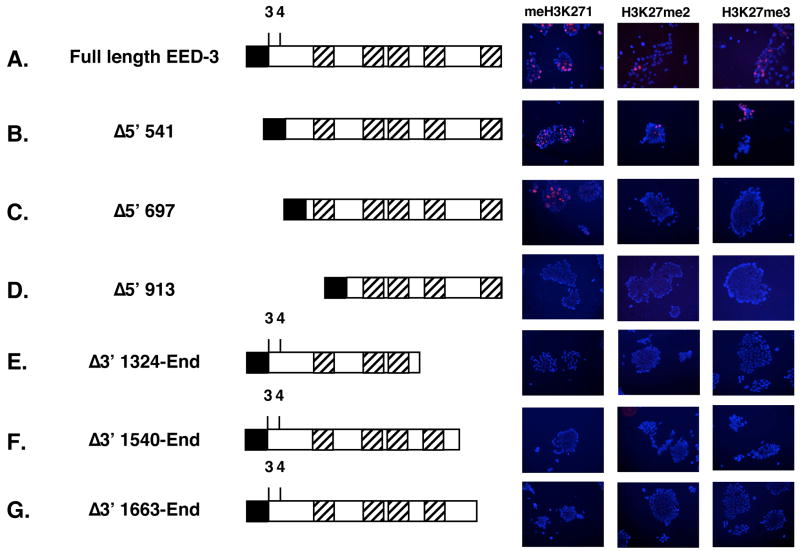
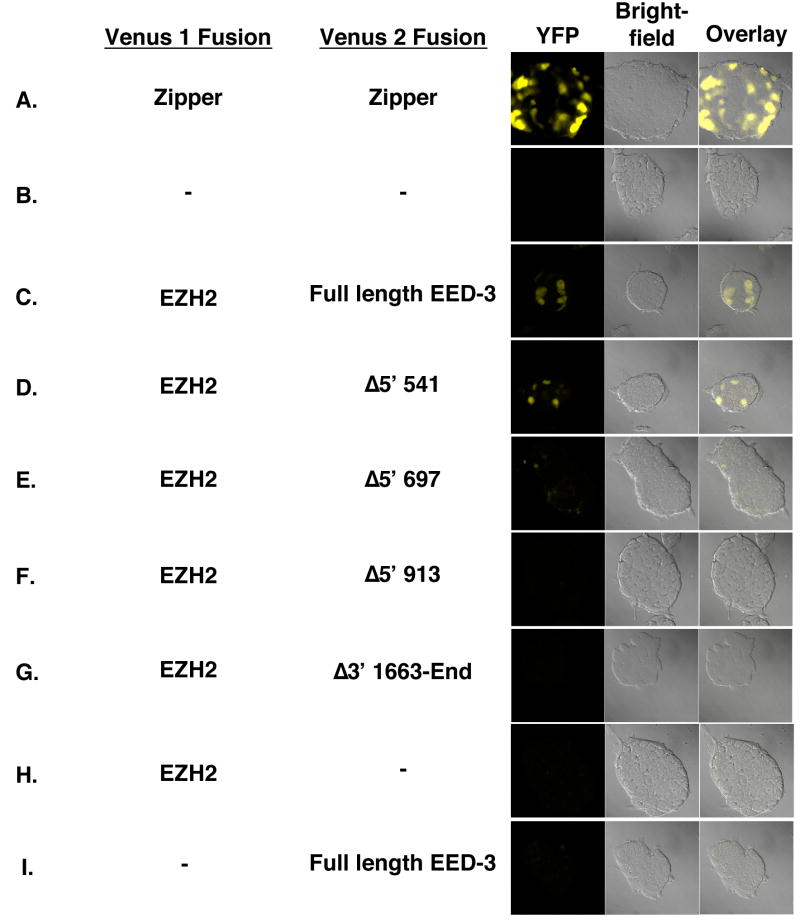
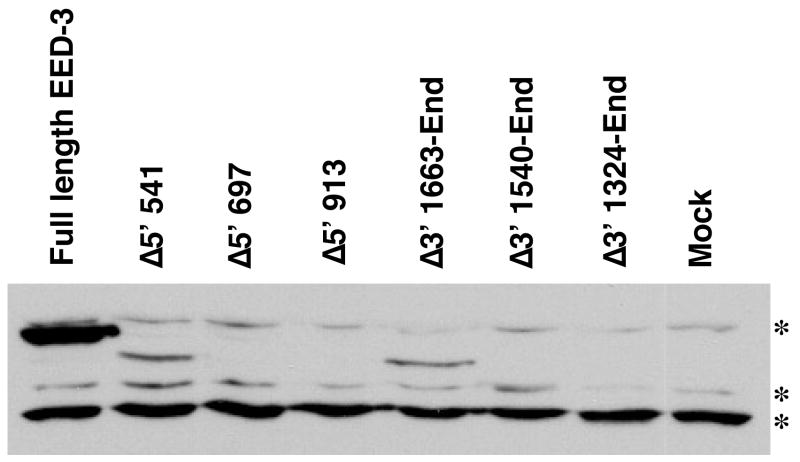
Polycomb group proteins represent a conserved family of developmental regulators that mediate heritable transcriptional silencing by modifying chromatin states. One Polycomb group complex, the PRC2 complex, is composed of several proteins, including the histone H3 lysine 27 (H3K27) methyltransferase enhancer of zeste homolog 2 and the WD-repeat protein embryonic ectoderm development (EED). Histone H3K27 can be monomethylated (H3K27me1), dimethylated (H3K27me2), or trimethylated (H3K27me3). However, it remains unclear what regulates the number of methyl groups added to H3K27 in a particular nucleosome. In mammalian cells, EED is present as four distinct isoforms, which are believed to be produced by utilizing four distinct, in-frame translation start sites in a common Eed mRNA. A mutation that disables all four EED isoforms produces defects in H3K27 methylation [Montgomery, N.D., Yee, D., Chen, A., Kalantry, S., Chamberlain, S.J., Otte, A.P. & Magnuson, T. (2005). The murine polycomb group protein Eed is required for global histone H3 lysine-27 methylation. Curr. Biol., 15, 942-947]. To assess the roles of individual EED isoforms in H3K27 methylation, we first characterized three of the four EED isoform start sites and then demonstrated that individual isoforms are not necessary for H3K27me1, H3K27me2, or H3K27me3. Instead, we show that the core WD-40 motifs and the histone-binding region of EED alone are sufficient for the generation of all three marks, demonstrating that EED isoforms do not control the number of methyl groups added to H3K27.
Figures
Similar articles
-
Binding of different histone marks differentially regulates the activity and specificity of polycomb repressive complex 2 (PRC2).Proc Natl Acad Sci U S A. 2010 Nov 9;107(45):19266-71. doi: 10.1073/pnas.1008937107. Epub 2010 Oct 25. Proc Natl Acad Sci U S A. 2010. PMID: 20974918 Free PMC article.
-
The murine polycomb group protein Eed is required for global histone H3 lysine-27 methylation.Curr Biol. 2005 May 24;15(10):942-7. doi: 10.1016/j.cub.2005.04.051. Curr Biol. 2005. PMID: 15916951
-
Role of the polycomb protein EED in the propagation of repressive histone marks.Nature. 2009 Oct 8;461(7265):762-7. doi: 10.1038/nature08398. Epub 2009 Sep 20. Nature. 2009. PMID: 19767730 Free PMC article.
-
Inner workings and regulatory inputs that control Polycomb repressive complex 2.Chromosoma. 2012 Jun;121(3):221-34. doi: 10.1007/s00412-012-0361-1. Epub 2012 Feb 19. Chromosoma. 2012. PMID: 22349693 Free PMC article. Review.
-
Structural basis for PRC2 engagement with chromatin.Curr Opin Struct Biol. 2021 Apr;67:135-144. doi: 10.1016/j.sbi.2020.10.017. Epub 2020 Nov 21. Curr Opin Struct Biol. 2021. PMID: 33232890 Review.
Cited by
-
Diverse involvement of EZH2 in cancer epigenetics.Am J Transl Res. 2015 Feb 15;7(2):175-93. eCollection 2015. Am J Transl Res. 2015. PMID: 25901190 Free PMC article. Review.
-
Degradation of Polycomb Repressive Complex 2 with an EED-Targeted Bivalent Chemical Degrader.Cell Chem Biol. 2020 Jan 16;27(1):47-56.e15. doi: 10.1016/j.chembiol.2019.11.006. Epub 2019 Dec 9. Cell Chem Biol. 2020. PMID: 31831267 Free PMC article.
-
Epigenetic regulation of embryonic ectoderm development in stem cell differentiation and transformation during ontogenesis.Cell Prolif. 2023 Apr;56(4):e13413. doi: 10.1111/cpr.13413. Epub 2023 Feb 1. Cell Prolif. 2023. PMID: 36727213 Free PMC article. Review.
-
Phosphorylation of EZH2 by CDK1 and CDK2: a possible regulatory mechanism of transmission of the H3K27me3 epigenetic mark through cell divisions.Cell Cycle. 2011 Feb 15;10(4):579-83. doi: 10.4161/cc.10.4.14850. Epub 2011 Feb 15. Cell Cycle. 2011. PMID: 21278485 Free PMC article.
-
PINK1 regulates histone H3 trimethylation and gene expression by interaction with the polycomb protein EED/WAIT1.Proc Natl Acad Sci U S A. 2013 Sep 3;110(36):14729-34. doi: 10.1073/pnas.1216844110. Epub 2013 Aug 19. Proc Natl Acad Sci U S A. 2013. Retraction in: Proc Natl Acad Sci U S A. 2014 Nov 11;111(45):16225. doi: 10.1073/pnas.1419356111. PMID: 23959866 Free PMC article. Retracted.
References
-
- Montgomery ND, Yee D, Chen A, Kalantry S, Chamberlain SJ, Otte AP, Magnuson T. The murine polycomb group protein Eed is required for global histone H3 lysine-27 methylation. Curr Biol. 2005;15:942–7. - PubMed
-
- Plath K, Fang J, Mlynarczyk-Evans SK, Cao R, Worringer KA, Wang H, de la Cruz CC, Otte AP, Panning B, Zhang Y. Role of histone H3 lysine 27 methylation in X inactivation. Science. 2003;300:131–5. - PubMed
-
- Silva J, Mak W, Zvetkova I, Appanah R, Nesterova TB, Webster Z, Peters AH, Jenuwein T, Otte AP, Brockdorff N. Establishment of histone h3 methylation on the inactive X chromosome requires transient recruitment of Eed-Enx1 polycomb group complexes. Dev Cell. 2003;4:481–95. - PubMed
-
- Gebuhr TC, Bultman SJ, Magnuson T. Pc-G/trx-G and the SWI/SNF connection: developmental gene regulation through chromatin remodeling. Genesis. 2000;26:189–97. - PubMed
-
- Lewis A, Mitsuya K, Umlauf D, Smith P, Dean W, Walter J, Higgins M, Feil R, Reik W. Imprinting on distal chromosome 7 in the placenta involves repressive histone methylation independent of DNA methylation. Nat Genet. 2004;36:1291–5. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
