

Regulation of tradeoffs between plant defenses against pathogens with different lifestyles
- PMID: 17998535
- PMCID: PMC2141864
- DOI: 10.1073/pnas.0708139104
Regulation of tradeoffs between plant defenses against pathogens with different lifestyles
Abstract
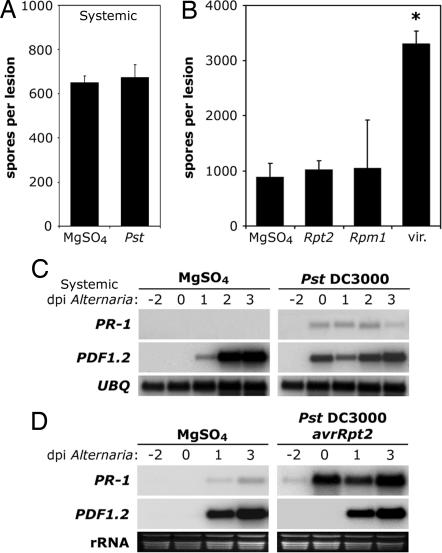
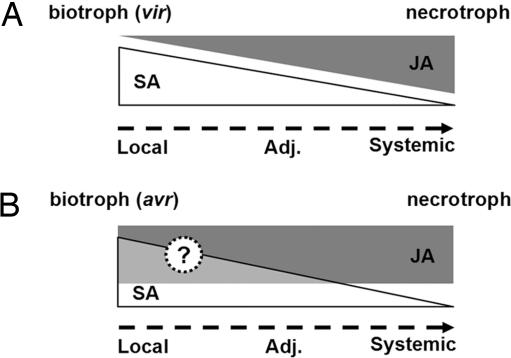
Plants activate distinct defense responses depending on the lifestyle of the attacker encountered. In these responses, salicylic acid (SA) and jasmonic acid (JA) play important signaling roles. SA induces defense against biotrophic pathogens that feed and reproduce on live host cells, whereas JA activates defense against necrotrophic pathogens that kill host cells for nutrition and reproduction. Cross-talk between these defense signaling pathways has been shown to optimize the response against a single attacker. However, its role in defense against multiple pathogens with distinct lifestyles is unknown. Here we show that infection with biotrophic Pseudomonas syringae, which induces SA-mediated defense, rendered plants more susceptible to the necrotrophic pathogen Alternaria brassicicola by suppression of the JA signaling pathway. This process was partly dependent on the cross-talk modulator NPR1. Surprisingly, this tradeoff was restricted to tissues adjacent to the site of initial infection; A. brassicicola infection in systemic tissue was not affected. Even more surprisingly, tradeoff occurred only with the virulent Pseudomonas strain. Avirulent strains that induced programmed cell death (PCD), an effective plant-resistance mechanism against biotrophs, did not cause suppression of JA-dependent defense. This result might be advantageous to the plant by preventing necrotrophic pathogen growth in tissues undergoing PCD. Our findings show that plants tightly control cross-talk between SA- and JA-dependent defenses in a previously unrecognized spatial and pathogen type-specific fashion. This process allows them to prevent unfavorable signal interactions and maximize their ability to concomitantly fend off multiple pathogens.
Conflict of interest statement
The authors declare no conflict of interest.
Figures


Similar articles
-
Ethylene and jasmonic acid signaling affect the NPR1-independent expression of defense genes without impacting resistance to Pseudomonas syringae and Peronospora parasitica in the Arabidopsis ssi1 mutant.Mol Plant Microbe Interact. 2003 Jul;16(7):588-99. doi: 10.1094/MPMI.2003.16.7.588. Mol Plant Microbe Interact. 2003. PMID: 12848424
-
Ethylene modulates the role of NONEXPRESSOR OF PATHOGENESIS-RELATED GENES1 in cross talk between salicylate and jasmonate signaling.Plant Physiol. 2009 Apr;149(4):1797-809. doi: 10.1104/pp.108.133926. Epub 2009 Jan 28. Plant Physiol. 2009. PMID: 19176718 Free PMC article.
-
AtOZF1 positively regulates JA signaling and SA-JA cross-talk in Arabidopsis thaliana.J Biosci. 2022;47:8. J Biosci. 2022. PMID: 35092410
-
Fine-Tuning Plant Defence Signalling: Salicylate versus Jasmonate.Plant Biol (Stuttg). 2006 Jan;8(1):1-10. doi: 10.1055/s-2005-872705. Plant Biol (Stuttg). 2006. PMID: 16435264 Review.
-
Signaling Crosstalk between Salicylic Acid and Ethylene/Jasmonate in Plant Defense: Do We Understand What They Are Whispering?Int J Mol Sci. 2019 Feb 4;20(3):671. doi: 10.3390/ijms20030671. Int J Mol Sci. 2019. PMID: 30720746 Free PMC article. Review.
Cited by
-
Salicylic acid: an old hormone up to new tricks.Mol Plant Pathol. 2013 Aug;14(6):623-34. doi: 10.1111/mpp.12035. Epub 2013 Apr 28. Mol Plant Pathol. 2013. PMID: 23621321 Free PMC article. Review.
-
The RNA silencing enzyme RNA polymerase v is required for plant immunity.PLoS Genet. 2011 Dec;7(12):e1002434. doi: 10.1371/journal.pgen.1002434. Epub 2011 Dec 29. PLoS Genet. 2011. PMID: 22242006 Free PMC article.
-
RING-Type E3 Ubiquitin Ligases AtRDUF1 and AtRDUF2 Positively Regulate the Expression of PR1 Gene and Pattern-Triggered Immunity.Int J Mol Sci. 2022 Nov 22;23(23):14525. doi: 10.3390/ijms232314525. Int J Mol Sci. 2022. PMID: 36498851 Free PMC article.
-
A Case of Plant Vaccination: Enhancement of Plant Immunity against Verticillium dahliae by Necrotized Spores of the Pathogen.Plants (Basel). 2022 Jun 26;11(13):1691. doi: 10.3390/plants11131691. Plants (Basel). 2022. PMID: 35807643 Free PMC article.
-
Water Deficit Timing Differentially Affects Physiological Responses of Grapevines Infected with Lasiodiplodia theobromae.Plants (Basel). 2022 Jul 28;11(15):1961. doi: 10.3390/plants11151961. Plants (Basel). 2022. PMID: 35956441 Free PMC article.
References
-
- Hancock JG, Huisman OC. Annu Rev Phytopathol. 1981;19:309–331.
-
- Glazebrook J. Annu Rev Phytopathol. 2005;43:205–227. - PubMed
-
- Malamy J, Carr JP, Klessig DF, Raskin I. Science. 1990;250:1004–1006. - PubMed
-
- Métraux J-P, Signer H, Ryals J, Ward E, Wyss-Benz M, Gaudin J, Raschdorf K, Schmid E, Blum W, Inverardi B. Science. 1990;250:1004–1006. - PubMed
-
- Maleck K, Levine A, Eulgem T, Morgan A, Schmid J, Lawton KA, Dangl JL, Dietrich RA. Nat Genet. 2000;26:403–410. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
