

Tissue interactions in the developing chick diencephalon
- PMID: 17999760
- PMCID: PMC2217525
- DOI: 10.1186/1749-8104-2-25
Tissue interactions in the developing chick diencephalon
Abstract
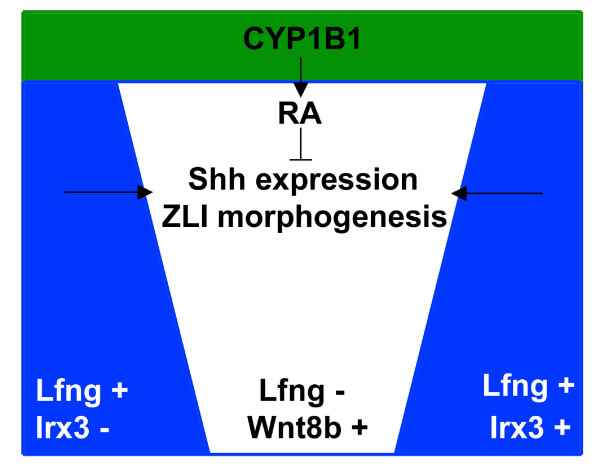
Background: The developing vertebrate brain is patterned first by global signalling gradients that define crude anteroposterior and dorsoventral coordinates, and subsequently by local signalling centres (organisers) that refine cell fate assignment within pre-patterned regions. The interface between the prethalamus and the thalamus, the zona limitans intrathalamica (ZLI), is one such local signalling centre that is essential for the establishment of these major diencephalic subdivisions by secreting the signalling factor Sonic hedgehog. Various models for ZLI formation have been proposed, but a thorough understanding of how this important local organiser is established is lacking.
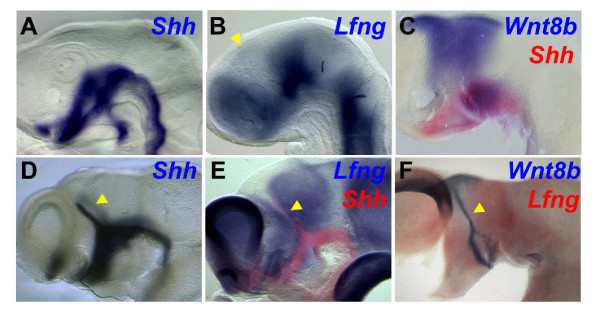
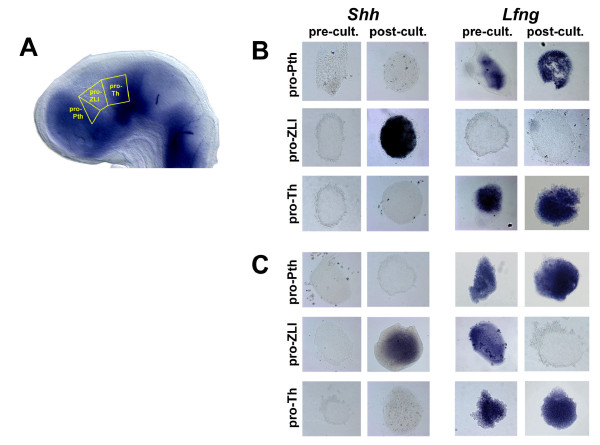
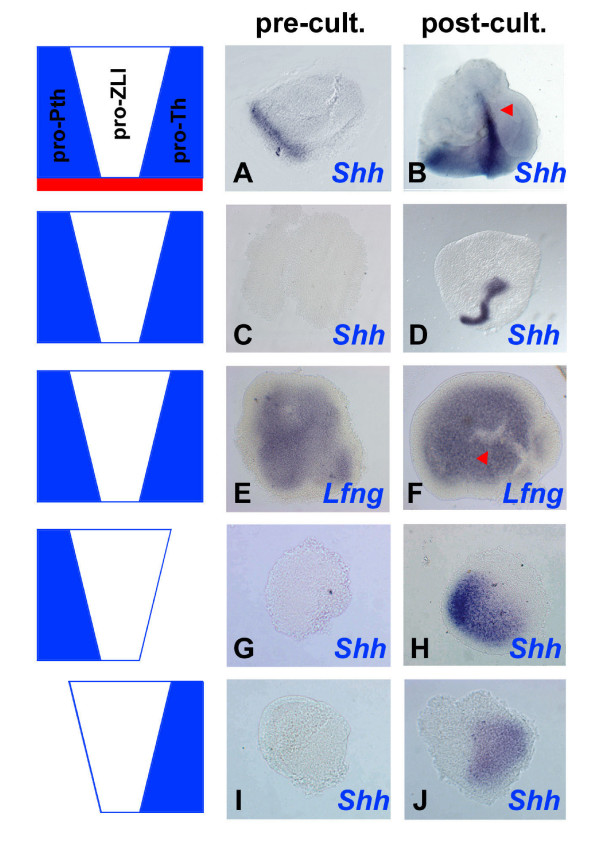
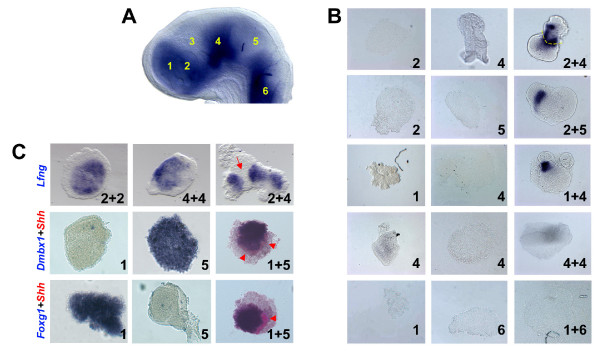
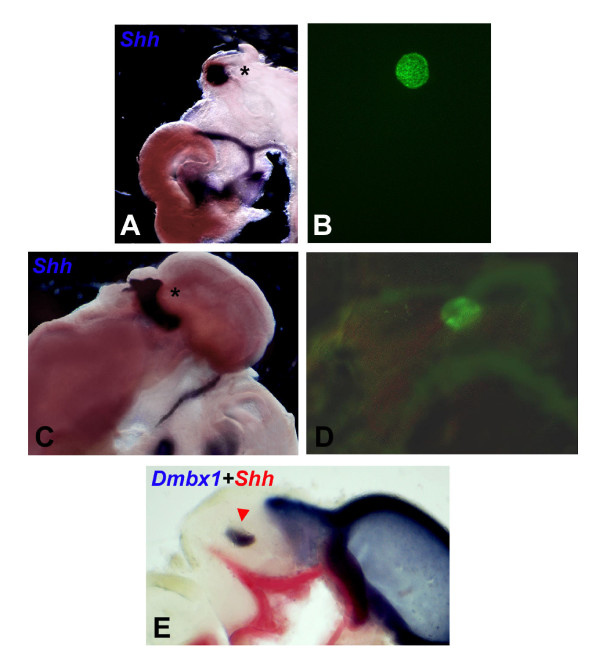
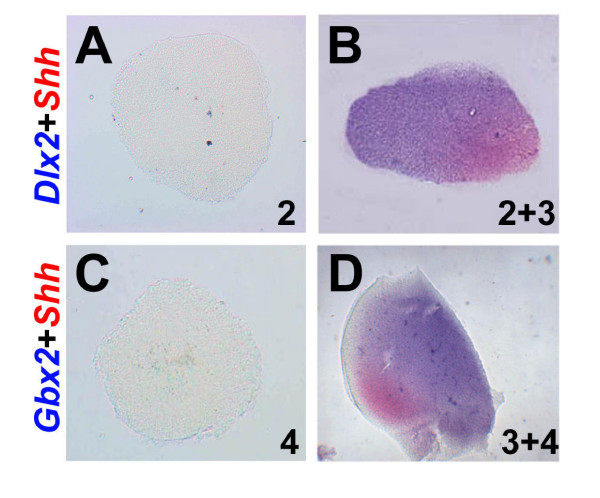
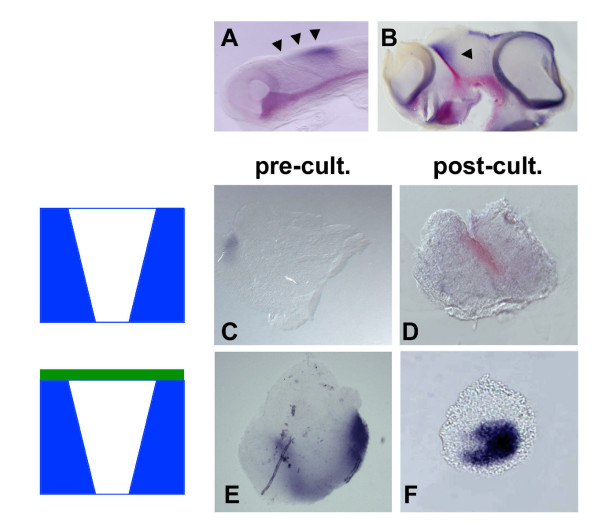
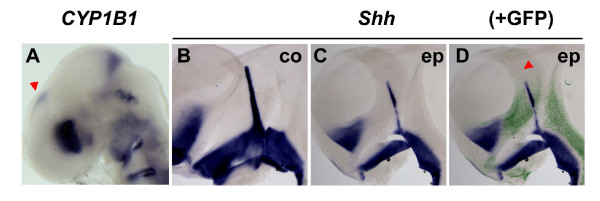
Results: Here, we describe tissue explant experiments in chick embryos aimed at characterising the roles of different forebrain areas in ZLI formation. We found that: the ZLI becomes specified unexpectedly early; flanking regions are required for its characteristic morphogenesis; ZLI induction can occur independently from ventral tissues; interaction between any prechordal and epichordal neuroepithelial tissue anterior to the midbrain-hindbrain boundary is able to generate a ZLI; and signals from the dorsal diencephalon antagonise ZLI formation. We further show that a localised source of retinoic acid in the dorsal diencephalon is a likely candidate to mediate this inhibitory signal.
Conclusion: Our results are consistent with a model where planar, rather than vertical, signals position the ZLI at early stages of neural development and they implicate retinoic acid as a novel molecular cue that determines its dorsoventral extent.
Figures
Similar articles
-
Hedgehog signalling from the zona limitans intrathalamica orchestrates patterning of the zebrafish diencephalon.Development. 2006 Mar;133(5):855-64. doi: 10.1242/dev.02248. Epub 2006 Feb 1. Development. 2006. PMID: 16452095
-
Sonic hedgehog from the basal plate and the zona limitans intrathalamica exhibits differential activity on diencephalic molecular regionalization and nuclear structure.Neuroscience. 2006 Nov 17;143(1):129-40. doi: 10.1016/j.neuroscience.2006.08.032. Epub 2006 Oct 10. Neuroscience. 2006. PMID: 17045408
-
Analysis of Lrrn1 expression and its relationship to neuromeric boundaries during chick neural development.Neural Dev. 2007 Oct 31;2:22. doi: 10.1186/1749-8104-2-22. Neural Dev. 2007. PMID: 17973992 Free PMC article.
-
Patterning the developing diencephalon.Brain Res Rev. 2007 Jan;53(1):17-26. doi: 10.1016/j.brainresrev.2006.06.004. Epub 2006 Jul 31. Brain Res Rev. 2007. PMID: 16876871 Review.
-
The role of Fgf8 in telencephalic and diencephalic patterning.Semin Cell Dev Biol. 2009 Aug;20(6):719-25. doi: 10.1016/j.semcdb.2009.04.002. Epub 2009 Apr 10. Semin Cell Dev Biol. 2009. PMID: 19596327 Review.
Cited by
-
Hindbrain boundaries as niches of neural progenitor and stem cells regulated by the extracellular matrix proteoglycan chondroitin sulphate.Development. 2024 Feb 15;151(4):dev201934. doi: 10.1242/dev.201934. Epub 2024 Feb 13. Development. 2024. PMID: 38251863 Free PMC article.
-
The Tale of the Three Brothers - Shh, Wnt, and Fgf during Development of the Thalamus.Front Neurosci. 2012 May 28;6:76. doi: 10.3389/fnins.2012.00076. eCollection 2012. Front Neurosci. 2012. PMID: 22654733 Free PMC article.
-
Diencephalic Size Is Restricted by a Novel Interplay Between GCN5 Acetyltransferase Activity and Retinoic Acid Signaling.J Neurosci. 2017 Mar 8;37(10):2565-2579. doi: 10.1523/JNEUROSCI.2121-16.2017. Epub 2017 Feb 2. J Neurosci. 2017. PMID: 28154153 Free PMC article.
-
Expression of hindbrain boundary markers is regulated by FGF3.Biol Open. 2012 Feb 15;1(2):67-74. doi: 10.1242/bio.2011032. Epub 2011 Nov 1. Biol Open. 2012. PMID: 23213398 Free PMC article.
-
Wnt signal specifies the intrathalamic limit and its organizer properties by regulating Shh induction in the alar plate.J Neurosci. 2013 Feb 27;33(9):3967-80. doi: 10.1523/JNEUROSCI.0726-12.2013. J Neurosci. 2013. PMID: 23447606 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
